- Обмен фосфолипидов и стероидов

Содержание
- 2. Обмен холестерина циклический ненасыщенный спирт, нерастворим в воде, в основе холестерина лежит кольцо циклопентанпергидрофенантрена.
- 3. Холестерин содержится в коре надпочечников, мозге, нервной ткани, мышцах, соединительной ткани, жировой ткани.
- 4. Биологическая роль холестерина синтез желчных кислот, синтез половых гормонов, синтез кортикостероидов, синтез витамина D3, входит в
- 5. Холестерин – предшественник стероидных гормонов
- 6. 40% - холестерина поступает с пищей, 60% - синтезируется в организме. Пищевые источники холестерина: печень, яйца,
- 7. Синтез холестерина осуществляется в эндоплазматическом ретикулуме, цитозоле печени (80%), коже, стенке тонкой кишки.
- 8. Синтез холестерина включает 35 реакций, идёт в 3 стадии: образование из ацетил-КоА мевалоновой кислоты, образование из
- 9. Ацетил-КоА-ацетил трансфераза Ацетоацетил-КоА
- 10. ОМГ- КоА- синтаза β−Окси-β−метилглутарил-КоА Ацетоацетил-КоА H2O
- 11. β−Окси-β−метилглутарил-КоА ОМГ- КоА- редуктаза + 2 НАДФН + 2Н+ Мевалоновая кислота + 2 НАДФ+ +HS-KoA
- 12. Мевалоновая кислота Сквален Сквален Ланостерин Ланостерин (С30) Холестерин (С27)
- 13. Синтез холестерина
- 17. Регуляция синтеза холестерина осуществляется по принципу обратной связи: холестерин угнетает синтез фермента ОМГ-редуктазы. Если содержание холестерина
- 18. Скорость обновления холестерина высока в надпочечниках и печени, низкая в мозге.
- 19. Транспорт холестерина холестерин из пищи проникает в стенку сосуда и мембраны клеток с ЛПНП, а удаляется
- 20. Формы холестерина внутриклеточный (метаболический), мембранный (структурный), внеклеточный (транспортный). В плазме человека холестерин находится в составе ЛП
- 21. Индекс атерогенности у новорожденных 1, у лиц 30 лет менее 3, у больных атеросклерозом 5-6.
- 22. Мембранный холестерин Количество холестерина, оседающее в мембранах, зависит от: активности специфических мембранных рецепторов холестерина, соотношения в
- 23. Мембранный холестерин В мембранах холестерин оказывает разжижающее и конденсирующее действие. Любая клетка не любит свободный холестерин,
- 24. Эфиры холестерина (внутриклеточный холестерин) запасная форма холестерина, 70 % холестерина находится в виде эфиров. При недостатке
- 25. Эффективный способ снижения уровня мембранного холестерина увеличение в пище количества фосфолипидов, содержащих полиненасыщенные жирные кислоты.
- 26. ЛХАТ внеклеточную эстерификацию холестерина осуществляет ЛХАТ, а внутриклеточную – АХАТ (ацилКоАхолестеролацилтрансфераза). ЛХАТ освобождает мембраны от избыточного
- 27. Окисление холестерина - единственный процесс необратимого его устранения из мембран и ЛП комплексов, - происходит в
- 28. Скорость окисления холестерина регулируется 7α-гидроксилазой. Фермент активируется холестерином, ингибируется – желчными кислотами. Половые гормоны и тироксин
- 29. Содержание общего холестерина 3,2 - 6,2 ммоль/л 70% эфиры холестерина, 30% - свободный холестерин, Содержание холестерина
- 30. Содержание холестерина более 5,2 ммоль/л - фактор риска атеросклероза. При концентрации 5,2 - 6,5 ммоль/л надо
- 31. Баланс холестерина в тканях Увеличение холестерина в тканях при: захвате ЛПНП рецепторами, захвате холестеролсодержащих ЛП без
- 32. Уменьшение холестерина при переходе холестерина из мембран в ЛПВП, эстерификации холестерина, окислении холестерина (использование холестерина для
- 33. Риск ИБС увеличивается при уровне холестерина 5,2 ммоль/л. Низкий уровень холестерина свидетельствует о патологии: анемии, гипертиреозе,
- 34. Кетоновые тела образуются из ацетил-КоА, синтезируются в печени. Содержание кетоновых тел в крови здоровых людей: 0,8
- 35. Синтез кетоновых тел Ацетил-КоА-ацетил трансфераза Ацетоацетил-КоА
- 36. ОМГ- КоА- синтаза β−Окси-β−метилглутарил-КоА Ацетоацетил-КоА H2O
- 37. β−Окси-β−метилглутарил-КоА Ацетоацетат ОМГ-КоА-лиаза
- 38. β-оксибутират- дегидрогеназа Ацетон β−Гидроксимасляная кислота СО2 НАДН + Н+ НАД+ Ацетоацетат
- 39. Источники синтеза кетоновых тел жирные кислоты, кетопластичные АМК. Избыток ацетил-КоА, высвобожденный при окислении жирных кислот и
- 40. Кетоновые тела – поставщики топлива для мышц, почек, мозга. Для мозга основным энергетическим субстратом являются глюкоза
- 41. + АТФ + АМФ + ФФн Ацетоацетил-КоА АцилКоА- синтетаза Использование ацетоуксусной кислоты
- 42. Использование ацетоуксусной кислоты Тиолаза Ацетоацетил-КоА
- 43. Регуляция Инсулин активирует синтез жира, тормозит образование ОМГ, ингибирует образование ацетоновых тел, Глюкагон активирует синтез ацетоновых
- 44. Кетонемия, кетонурия наблюдаются при сахарном диабете, голодании, длительной мышечной работе, токсикозе беременных, приёме пищи, богатой жирами.
- 45. В мозге новорожденных кетоновые тела потребляются в 3 раза интенсивнее, чем у взрослых. В раннем детстве
- 46. Склонность к кетозу повышена у детей в возрасте от 2 до 10 лет, так как снижена
- 47. Синтез фосфолипидов + ЦТФ Фосфатидная кислота ЦДФ-диацилглицерид + ФФн
- 48. Серин ЦДФ-диацилглицерид Фосфатидилсерин + ЦМФ
- 49. Фосфатидилсерин - СО2 ФП Фосфатидилэтаноламин
- 50. Фосфатидилэтаноламин Фосфатидилхолин S-аденозил- метионин
- 51. Фосфолипиды могут синтезироваться из готовых остатков (резервный путь) холин + АТФ Ё фосфохолин + АДФ фосфохолин
- 52. этаноламин + АТФ Ёфосфоэтаноламин + АДФ фосфоэтаноламин + ЦТФ Ё ЦДФ- этаноламин + ФФн ЦДФ- этаноламин
- 53. Функции фосфолипидов структурная (входят в состав мембран, мозга), участвуют в обмене холестерина лецитин + холестерин Ёлизолецитин+
- 54. При уменьшении молекулярного соотношения ФЛ:ХС менее 3:2 рекомендуются липотропные диеты, богатые ФЛ.
- 55. Жировая инфильтрация печени развивается при действии гепатотропных ядов, вирусов.
- 56. Липотропные вещества способны предотвращать жировую инфильтрацию печени холин, метионин, лецитин, казеин, инозит, липокаин, витамин В12, фолиевая
- 57. Механизм липотропного действия Липотропные вещества необходимы для синтеза ЛП (холин). Синтез ЛП – важный путь утилизации
- 59. Скачать презентацию
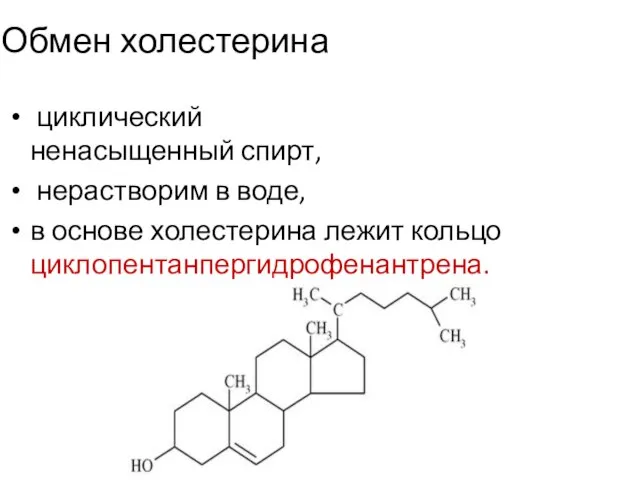
Обмен холестерина
циклический ненасыщенный спирт,
нерастворим в воде,
в основе холестерина лежит
Обмен холестерина
циклический ненасыщенный спирт,
нерастворим в воде,
в основе холестерина лежит

Холестерин содержится в
коре надпочечников,
мозге,
нервной ткани,
мышцах,
соединительной
Холестерин содержится в
коре надпочечников,
мозге,
нервной ткани,
мышцах,
соединительной

Биологическая роль холестерина
синтез желчных кислот,
синтез половых гормонов,
синтез кортикостероидов,
Биологическая роль холестерина
синтез желчных кислот,
синтез половых гормонов,
синтез кортикостероидов,
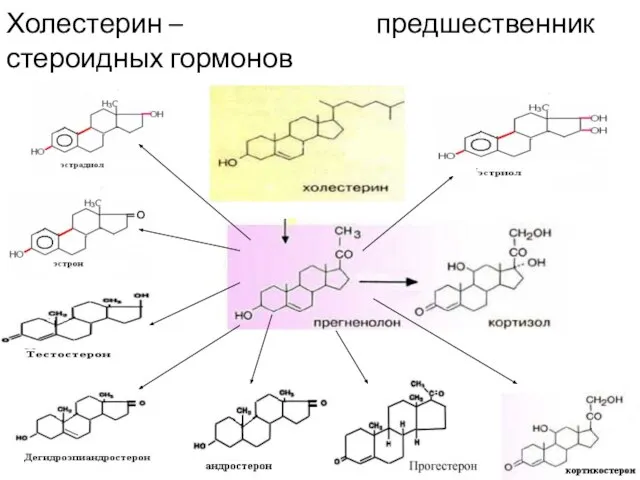
Холестерин – предшественник стероидных гормонов
Холестерин – предшественник стероидных гормонов

40% - холестерина поступает с пищей,
60% - синтезируется в организме.
Пищевые источники
40% - холестерина поступает с пищей,
60% - синтезируется в организме.
Пищевые источники

Синтез холестерина осуществляется в
эндоплазматическом ретикулуме,
цитозоле печени (80%),
коже,
Синтез холестерина осуществляется в
эндоплазматическом ретикулуме,
цитозоле печени (80%),
коже,
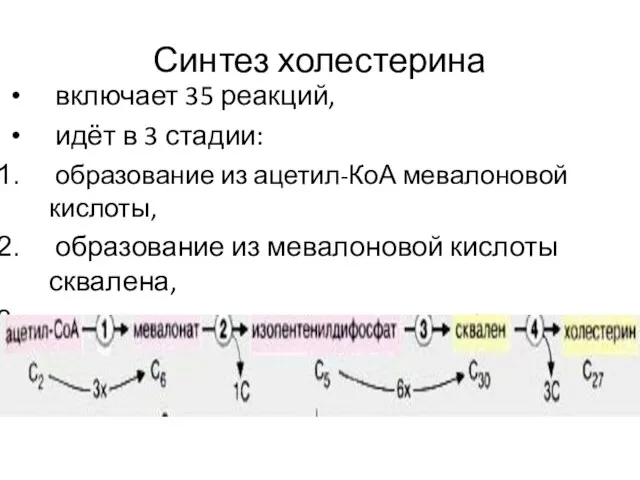
Синтез холестерина
включает 35 реакций,
идёт в 3 стадии:
образование из
Синтез холестерина
включает 35 реакций,
идёт в 3 стадии:
образование из
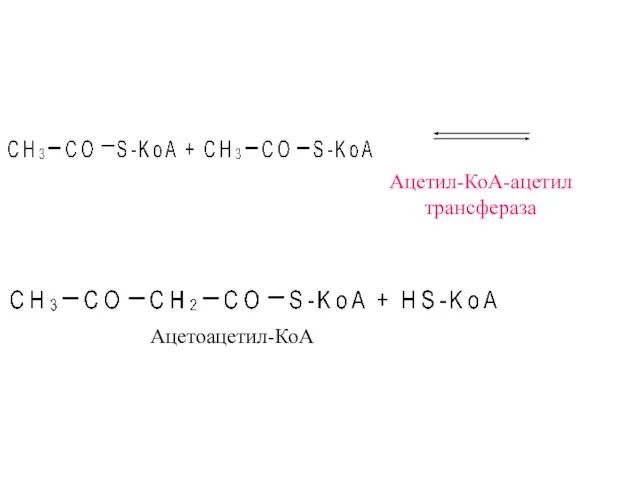
Ацетил-КоА-ацетил
трансфераза
Ацетоацетил-КоА
Ацетил-КоА-ацетил
трансфераза
Ацетоацетил-КоА
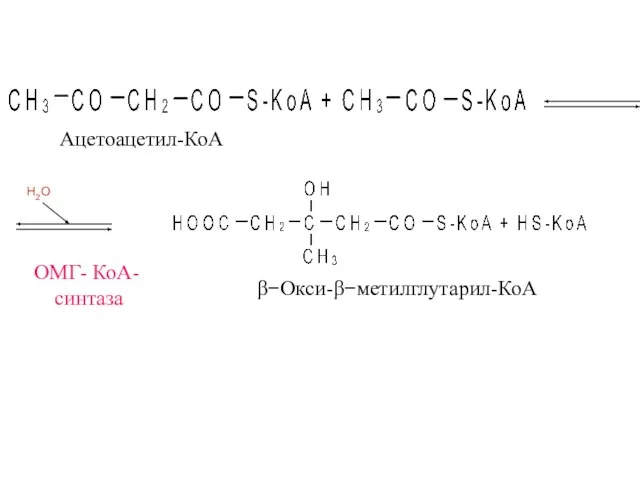
ОМГ- КоА-
синтаза
β−Окси-β−метилглутарил-КоА
Ацетоацетил-КоА
H2O
ОМГ- КоА-
синтаза
β−Окси-β−метилглутарил-КоА
Ацетоацетил-КоА
H2O
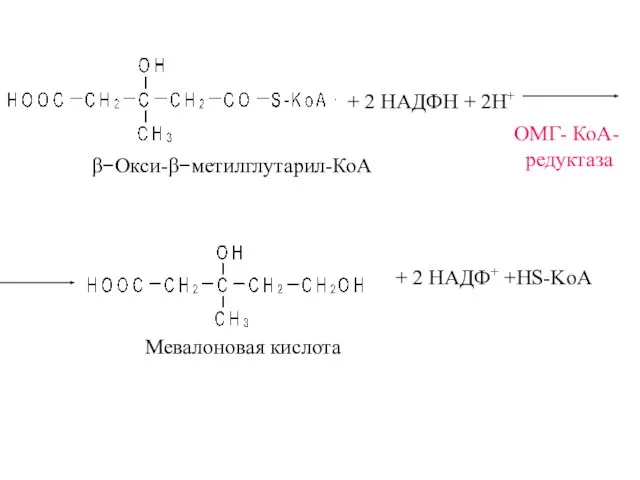
β−Окси-β−метилглутарил-КоА
ОМГ- КоА-
редуктаза
+ 2 НАДФН + 2Н+
Мевалоновая кислота
+ 2 НАДФ+ +HS-KoA
β−Окси-β−метилглутарил-КоА
ОМГ- КоА-
редуктаза
+ 2 НАДФН + 2Н+
Мевалоновая кислота
+ 2 НАДФ+ +HS-KoA
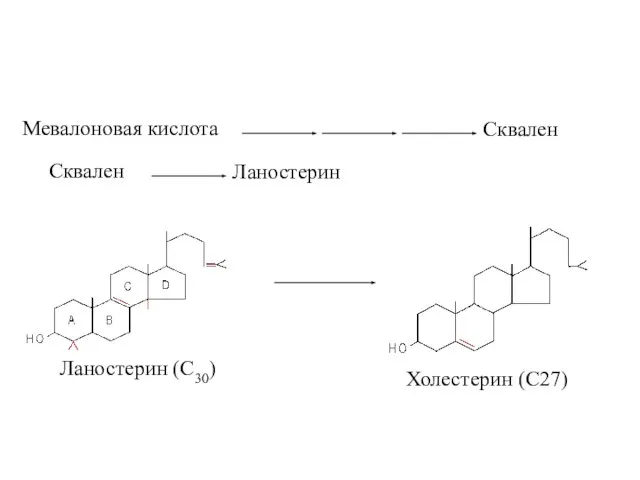
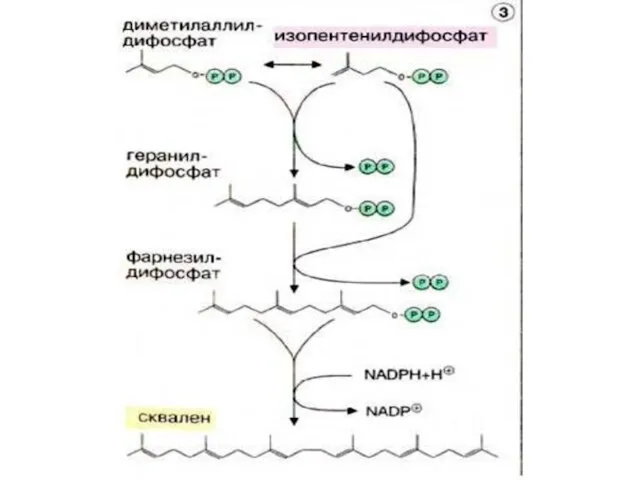
Мевалоновая кислота
Сквален
Сквален
Ланостерин
Ланостерин (С30)
Холестерин (С27)
Мевалоновая кислота
Сквален
Сквален
Ланостерин
Ланостерин (С30)
Холестерин (С27)
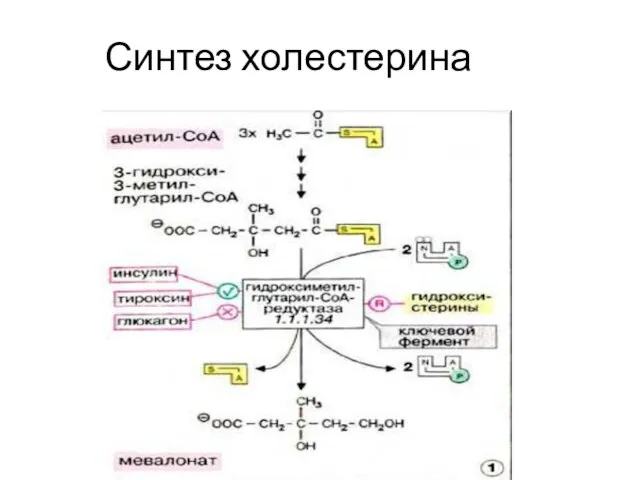
Синтез холестерина
Синтез холестерина



Регуляция синтеза холестерина
осуществляется по принципу обратной связи: холестерин угнетает синтез
Регуляция синтеза холестерина
осуществляется по принципу обратной связи: холестерин угнетает синтез

Скорость обновления холестерина
высока в надпочечниках и печени,
низкая в мозге.
Скорость обновления холестерина
высока в надпочечниках и печени,
низкая в мозге.

Транспорт холестерина
холестерин из пищи проникает в стенку сосуда и мембраны
Транспорт холестерина
холестерин из пищи проникает в стенку сосуда и мембраны

Формы холестерина
внутриклеточный (метаболический),
мембранный (структурный),
внеклеточный (транспортный).
В плазме человека холестерин
Формы холестерина
внутриклеточный (метаболический),
мембранный (структурный),
внеклеточный (транспортный).
В плазме человека холестерин

Индекс атерогенности
у новорожденных 1,
у лиц 30 лет менее 3,
Индекс атерогенности
у новорожденных 1,
у лиц 30 лет менее 3,

Мембранный
холестерин
Количество холестерина,
оседающее в мембранах, зависит от:
активности специфических мембранных
Мембранный
холестерин
Количество холестерина,
оседающее в мембранах, зависит от:
активности специфических мембранных

Мембранный холестерин
В мембранах холестерин оказывает разжижающее и конденсирующее действие.
Любая клетка не
Мембранный холестерин
В мембранах холестерин оказывает разжижающее и конденсирующее действие.
Любая клетка не

Эфиры холестерина (внутриклеточный холестерин)
запасная форма холестерина,
70 % холестерина находится
Эфиры холестерина (внутриклеточный холестерин)
запасная форма холестерина,
70 % холестерина находится

Эффективный способ снижения уровня мембранного холестерина
увеличение в пище количества фосфолипидов,
Эффективный способ снижения уровня мембранного холестерина
увеличение в пище количества фосфолипидов,

ЛХАТ
внеклеточную эстерификацию холестерина осуществляет ЛХАТ,
а внутриклеточную – АХАТ (ацилКоАхолестеролацилтрансфераза).
ЛХАТ
ЛХАТ
внеклеточную эстерификацию холестерина осуществляет ЛХАТ,
а внутриклеточную – АХАТ (ацилКоАхолестеролацилтрансфераза).
ЛХАТ

Окисление холестерина
- единственный процесс необратимого его устранения из мембран и ЛП
Окисление холестерина
- единственный процесс необратимого его устранения из мембран и ЛП

Скорость окисления холестерина
регулируется 7α-гидроксилазой.
Фермент активируется холестерином,
ингибируется – желчными
Скорость окисления холестерина
регулируется 7α-гидроксилазой.
Фермент активируется холестерином,
ингибируется – желчными

Содержание общего холестерина
3,2 - 6,2 ммоль/л
70% эфиры холестерина,
30% -
Содержание общего холестерина
3,2 - 6,2 ммоль/л
70% эфиры холестерина,
30% -

Содержание холестерина
более 5,2 ммоль/л -
фактор риска
атеросклероза.
При концентрации 5,2 -
Содержание холестерина
более 5,2 ммоль/л -
фактор риска
атеросклероза.
При концентрации 5,2 -

Баланс холестерина в тканях
Увеличение холестерина в тканях при:
захвате ЛПНП
Баланс холестерина в тканях
Увеличение холестерина в тканях при:
захвате ЛПНП

Уменьшение холестерина при
переходе холестерина из мембран в ЛПВП,
эстерификации
Уменьшение холестерина при
переходе холестерина из мембран в ЛПВП,
эстерификации

Риск ИБС увеличивается при уровне холестерина 5,2 ммоль/л.
Низкий уровень холестерина
Низкий уровень холестерина
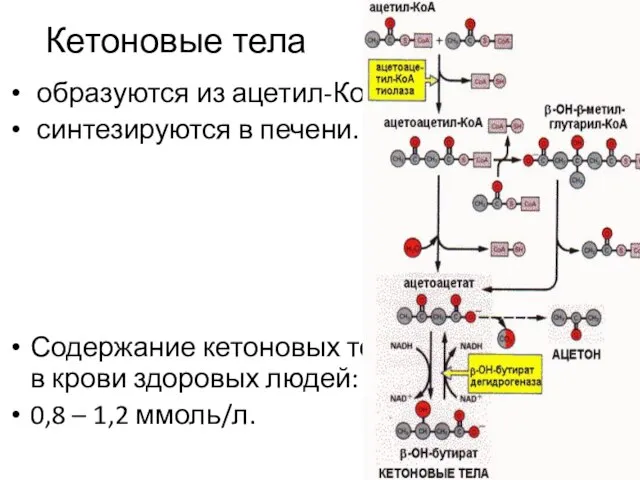
Кетоновые тела
образуются из ацетил-КоА,
синтезируются в печени.
Содержание кетоновых
Кетоновые тела
образуются из ацетил-КоА,
синтезируются в печени.
Содержание кетоновых
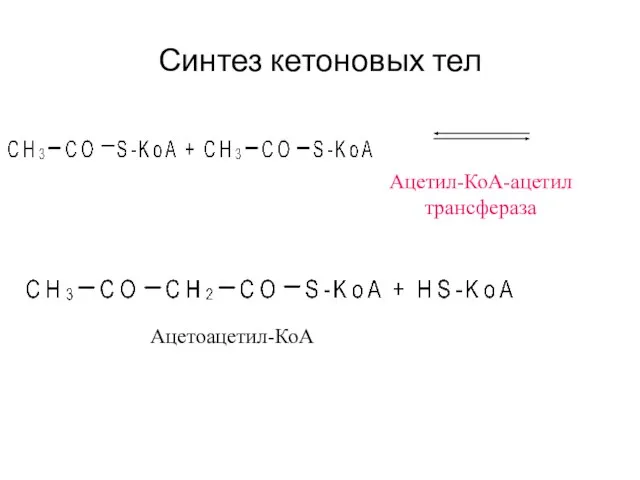
Синтез кетоновых тел
Ацетил-КоА-ацетил
трансфераза
Ацетоацетил-КоА
Синтез кетоновых тел
Ацетил-КоА-ацетил
трансфераза
Ацетоацетил-КоА
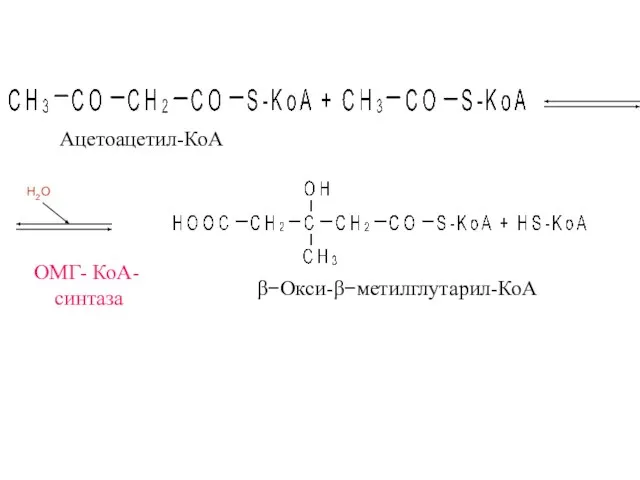
ОМГ- КоА-
синтаза
β−Окси-β−метилглутарил-КоА
Ацетоацетил-КоА
H2O
ОМГ- КоА-
синтаза
β−Окси-β−метилглутарил-КоА
Ацетоацетил-КоА
H2O
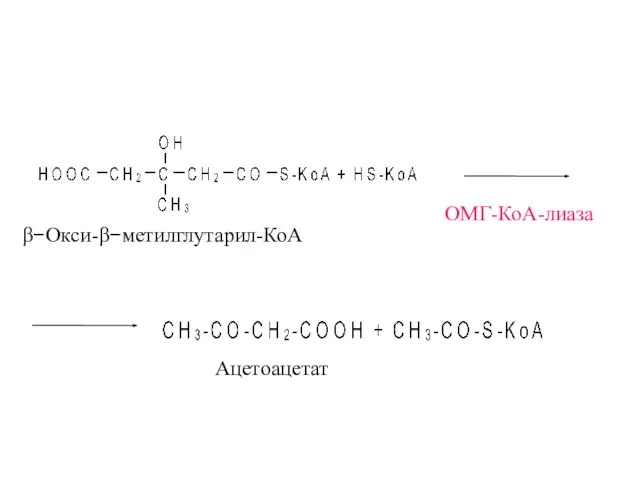
β−Окси-β−метилглутарил-КоА
Ацетоацетат
ОМГ-КоА-лиаза
β−Окси-β−метилглутарил-КоА
Ацетоацетат
ОМГ-КоА-лиаза
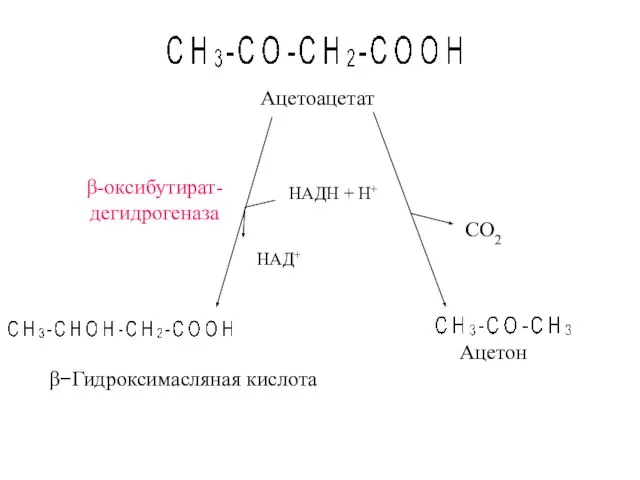
β-оксибутират-
дегидрогеназа
Ацетон
β−Гидроксимасляная кислота
СО2
НАДН + Н+
НАД+
Ацетоацетат
β-оксибутират-
дегидрогеназа
Ацетон
β−Гидроксимасляная кислота
СО2
НАДН + Н+
НАД+
Ацетоацетат

Источники синтеза кетоновых тел
жирные кислоты,
кетопластичные АМК.
Избыток ацетил-КоА,
Источники синтеза кетоновых тел
жирные кислоты,
кетопластичные АМК.
Избыток ацетил-КоА,

Кетоновые тела – поставщики топлива для
мышц,
почек,
мозга.
Для
Кетоновые тела – поставщики топлива для
мышц,
почек,
мозга.
Для
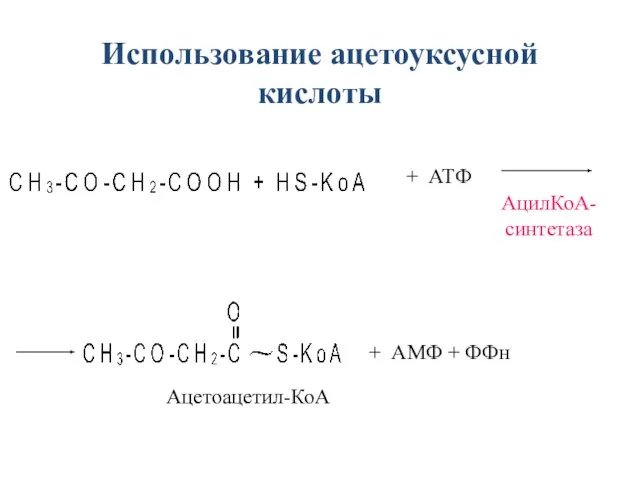
+ АТФ
+ АМФ + ФФн
Ацетоацетил-КоА
АцилКоА-
синтетаза
Использование ацетоуксусной кислоты
+ АТФ
+ АМФ + ФФн
Ацетоацетил-КоА
АцилКоА-
синтетаза
Использование ацетоуксусной кислоты
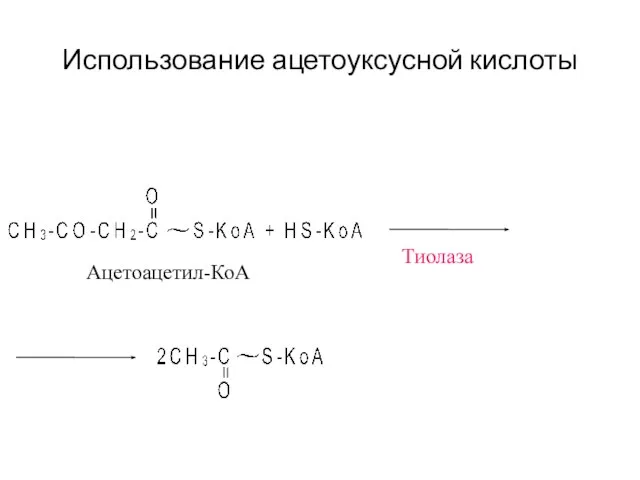
Использование ацетоуксусной кислоты
Тиолаза
Ацетоацетил-КоА
Использование ацетоуксусной кислоты
Тиолаза
Ацетоацетил-КоА

Регуляция
Инсулин
активирует синтез жира,
тормозит образование ОМГ,
ингибирует образование
Регуляция
Инсулин
активирует синтез жира,
тормозит образование ОМГ,
ингибирует образование

Кетонемия, кетонурия наблюдаются при
сахарном диабете,
голодании,
длительной мышечной работе,
токсикозе беременных,
Кетонемия, кетонурия наблюдаются при
сахарном диабете,
голодании,
длительной мышечной работе,
токсикозе беременных,

В мозге новорожденных
кетоновые тела потребляются в 3 раза интенсивнее, чем
В мозге новорожденных
кетоновые тела потребляются в 3 раза интенсивнее, чем

Склонность к кетозу
повышена у детей в возрасте от 2 до 10
Склонность к кетозу повышена у детей в возрасте от 2 до 10
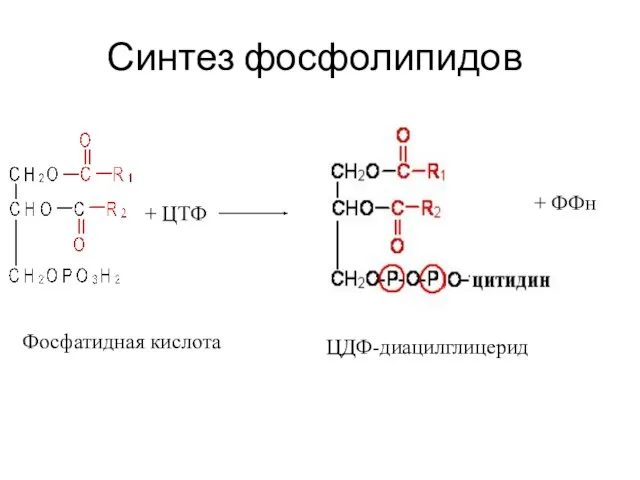
Синтез фосфолипидов
+ ЦТФ
Фосфатидная кислота
ЦДФ-диацилглицерид
+ ФФн
Синтез фосфолипидов
+ ЦТФ
Фосфатидная кислота
ЦДФ-диацилглицерид
+ ФФн
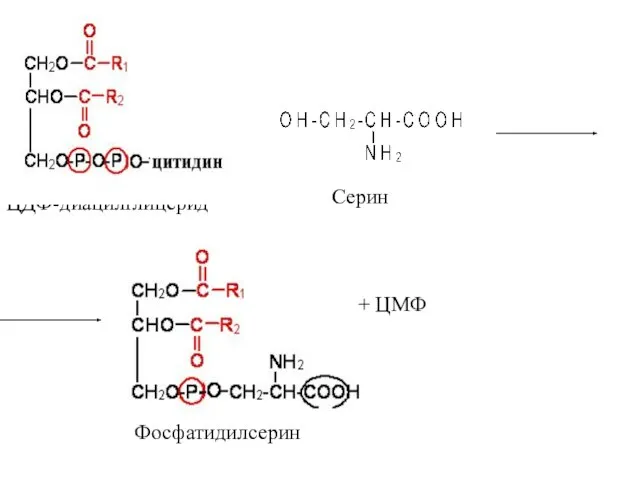
Серин
ЦДФ-диацилглицерид
Фосфатидилсерин
+ ЦМФ
Серин
ЦДФ-диацилглицерид
Фосфатидилсерин
+ ЦМФ
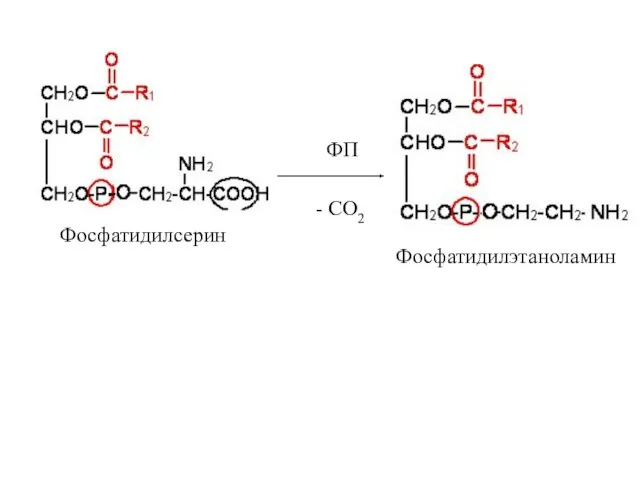
Фосфатидилсерин
- СО2
ФП
Фосфатидилэтаноламин
Фосфатидилсерин
- СО2
ФП
Фосфатидилэтаноламин
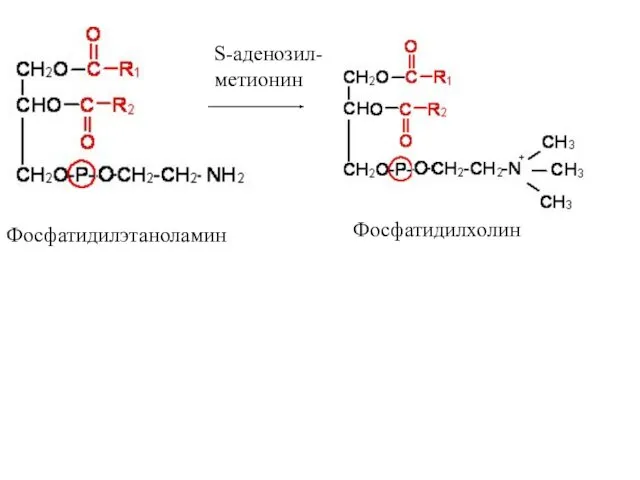
Фосфатидилэтаноламин
Фосфатидилхолин
S-аденозил-
метионин
Фосфатидилэтаноламин
Фосфатидилхолин
S-аденозил-
метионин

Фосфолипиды могут синтезироваться из готовых остатков (резервный путь)
холин + АТФ
Фосфолипиды могут синтезироваться из готовых остатков (резервный путь)
холин + АТФ

этаноламин + АТФ Ёфосфоэтаноламин + АДФ
фосфоэтаноламин + ЦТФ Ё ЦДФ-
этаноламин + АТФ Ёфосфоэтаноламин + АДФ
фосфоэтаноламин + ЦТФ Ё ЦДФ-

Функции фосфолипидов
структурная (входят в состав мембран, мозга),
участвуют в обмене
Функции фосфолипидов
структурная (входят в состав мембран, мозга),
участвуют в обмене

При уменьшении молекулярного соотношения ФЛ:ХС менее 3:2 рекомендуются липотропные диеты, богатые
При уменьшении молекулярного соотношения ФЛ:ХС менее 3:2 рекомендуются липотропные диеты, богатые

Жировая инфильтрация печени развивается при действии
гепатотропных ядов,
вирусов.
Жировая инфильтрация печени развивается при действии
гепатотропных ядов,
вирусов.

Липотропные вещества
способны предотвращать жировую инфильтрацию печени
холин,
метионин,
лецитин,
казеин,
Липотропные вещества
способны предотвращать жировую инфильтрацию печени
холин,
метионин,
лецитин,
казеин,

Механизм липотропного действия
Липотропные вещества необходимы для синтеза ЛП (холин). Синтез
Механизм липотропного действия
Липотропные вещества необходимы для синтеза ЛП (холин). Синтез
 Alkali metals
Alkali metals Получение, собирание, распознавание газов. (Практическая работа 2)
Получение, собирание, распознавание газов. (Практическая работа 2) Гетероциклические соединения
Гетероциклические соединения Воздух. Свойства воздуха
Воздух. Свойства воздуха Структура ячейки на примере атома железа
Структура ячейки на примере атома железа Пластмассы (пластики)
Пластмассы (пластики) Алкалоиды ряда изохинолина и пурина. (Лекция 10)
Алкалоиды ряда изохинолина и пурина. (Лекция 10) Жер қыртысының заттық және химиялық құрамы
Жер қыртысының заттық және химиялық құрамы Увеличение эффективности печи установки АТ
Увеличение эффективности печи установки АТ Каталітичні процеси нафтопереробки
Каталітичні процеси нафтопереробки Презентация по Химии "Йод" - скачать смотреть
Презентация по Химии "Йод" - скачать смотреть  Презентация по Химии "Польза жиров" - скачать смотреть
Презентация по Химии "Польза жиров" - скачать смотреть  Повторение 8 класса. Основные понятия химии
Повторение 8 класса. Основные понятия химии Добування кисню та вивчення його властивостей
Добування кисню та вивчення його властивостей Коллигативные свойства растворов
Коллигативные свойства растворов Углеводороды. Применение природных источников углеводородов и продуктов их переработки
Углеводороды. Применение природных источников углеводородов и продуктов их переработки Презентация по Химии "Нові підходи до відходів" - скачать смотреть бесплатно
Презентация по Химии "Нові підходи до відходів" - скачать смотреть бесплатно Реактор для гетерогенного катализа с движущим слоем катализатора
Реактор для гетерогенного катализа с движущим слоем катализатора 6 КАЗ Изохинолин туынд
6 КАЗ Изохинолин туынд Аттестационная работа. Образовательная программа элективного курса по химии Химия и мы
Аттестационная работа. Образовательная программа элективного курса по химии Химия и мы Электродные потенциалы
Электродные потенциалы Задачи в школьном курсе химии
Задачи в школьном курсе химии Презентация по химии Нуклеофильное замещение карбоновых кислот
Презентация по химии Нуклеофильное замещение карбоновых кислот Транскрипция. Биосинтез РНК. (Лекция 7)
Транскрипция. Биосинтез РНК. (Лекция 7) Ковалентная химическая связь. 11 класс
Ковалентная химическая связь. 11 класс Химия и здоровье человека
Химия и здоровье человека Буферные растворы
Буферные растворы ГИА. Вопрос А9. Химические свойства простых веществ: металлов и неметаллов
ГИА. Вопрос А9. Химические свойства простых веществ: металлов и неметаллов