- Репликация ДНК и РНК

Содержание
- 2. ФУНКЦИИ НУКЛЕИНОВЫХ КИСЛОТ КАК ГЕНЕТИЧЕСКОГО МАТЕРИАЛА Хранение, передача и реализация генетической информации Хранение – содержание информации
- 3. Центральная догма молекулярной биологии Фрэнсис Крик (Francis Harry Compton Crick) 1916 - 2004 ДНК РНК Белок
- 4. ДНК Хранение и передача нуклеиновыми кислотами генетической информации в неизменном виде
- 5. Репликация Удвоение ДНК в ходе деления клетки 2n 4n 2n 2n
- 6. Репликация – удвоение ДНК Репликация +
- 7. Модели репликации Репликация Консервативный механизм Полуконсервативный механизм Дисперсивный механизм + + +
- 8. Полуконсервативный механизм репликации Репликация Мэтью Стэнли Мезенсон Matthew Stanley Meselson 1930 г.р. Франклин Уильям Сталь Franklin
- 9. Эксперимент Мезельсона-Сталя (PNAS.-1958.-44:671-682) Репликация 14N - 99,635 % и 15N - 0,365 % 15NH4Cl E.coli E.coli
- 10. Репликация Формирование градиента плотности CsCl (исходная концентрация 7.75 М) и образование узкой зоны ДНК в области
- 11. Репликация Зоны ДНК, очищенной из E.coli после выращивания с 15NH4Cl и c 14NH4Cl. Разница в плавучих
- 12. Репликация ДНК из E.coli после выращивания с 15NH4Cl ДНК из E.coli после выращивания с 14NH4Cl в
- 13. Репликация Что доказывает эксперимент Мезельсона-Сталя? 0 1 2 3 Дисперсивный механизм 15N 15N 14N 15N 14N
- 14. Репликация Точка начала репликации и репликационные вилки ori Репликация ДНК бактерий + Репликационная вилка
- 15. 13-19 мин Репликация Точка начала репликации и репликационные вилки ori ori Эксперимент с меченными нуклеотидами разной
- 16. Репликация Репликация кольцевой ДНК механизм «катящегося кольца» Дочерняя ДНК
- 17. Репликация Репликация ДНК эукариот множественные точки начала репликации ori ori ori
- 18. Небольшое отступление Кольцевая и псевдокольцевая ДНК Надцарство прокариот (3.5 млрд. лет) Надцарство эукариот (1.5-2 млрд. лет)
- 20. 1. Оксидоредуктазы 2. Трансферазы 3. Гидролазы 4. Лиазы 5. Изомеразы 6. Лигазы Небольшое отступление A(окисл) +
- 21. Репликация ДНК-полимераза (трансфераза) α-[32P]-дезоксирибонуклеотиды α-[32P]-дАТФ α-[32P]-дГТФ α-[32P]-дЦТФ α-[32P]-дТТФ + экстракт E.coli Очистка ДНК Измерение радиоактивности ДНК
- 22. Очистка белков Освновной принцип – обогащение раствора требуемым белком Избирательное осаждение
- 23. Очистка белков Избирательное осаждение белков Изоэлектрическая точка и зависимость растворимости от pH -H+ +H+ -H+ +H+
- 24. Избирательное осаждение белков Изоэлектрическая точка и зависимость растворимости от pH pH 6 pH > 8 –
- 25. Избирательное осаждение белков Изоэлектрическая точка и зависимость растворимости от pH + + + + + +
- 26. Избирательное осаждение белков Высаливание В водном растворе белки связывают часть молекул воды. При замораживании незамороженными (не
- 27. Избирательное осаждение белков Высаливание Гидрофобные участки на поверхности белка Гидратная оболочка белка Белки, имеющие гидратную оболочку,
- 28. Избирательное осаждение белков Высаливание Очистка белков
- 29. Избирательное осаждение белков Денатурация (удаление из смеси ненужных белков) pH (сильный сдвиг) Температура Органические растворители Денатурация
- 30. Хроматография (гель-фильтрация) OD Очистка белков
- 31. Репликация ДНК-полимераза Фракция, обогащенная белком (ДНК-полимеразой) Экстракт ткани Добавляем фракцию, обогащенную белком Добавляем радиоактивно меченные дезоксирибонуклеотиды
- 32. Удаляем нуклеотиды Удаляем нуклеотиды Добавляем фракцию, обогащенную белком Добавляем радиоактивно меченные рибонуклеотиды Инкубируем Радиоактивность в растворе
- 33. Добавляем фракцию, обогащенную белком Добавляем радиоактивно меченные рибонуклеотиды Инкубируем Следовательно, для синтеза ДНК нужна полимерная ДНК
- 34. Репликация ДНК-полимераза (трансфераза) (дNМФ)n + дNТФ (дNМФ)n+1 + PPi ДНК-полимераза ДНК-матрица ДНК-матрица Дочерняя ДНК
- 36. ДНК-полимераза требует наличия матрицы (материнской ДНК). ДНК-полимераза присоединяет дезоксирибонуклеозидмонофосфат в соответствии с правилом комплементарности относительно нуклеотида
- 37. P G 5’ 3’ 5’-pApCpGpTpC-3’ P P A C P C P T 3’ 5’ Первичная
- 38. ДНК-полимераза работает только на одноцепочечной ДНК Репликация ДНК-полимераза 5’ 5’ 3’ 3’ Для работы ДНК-полимеразы требуется
- 39. ДНК-полимераза не может начать синтез ДНК с первого нуклеотида, она присоединяет дезоксирибонуклеотидмонофосфат только к уже существующей
- 40. Репликация ДНК-полимераза ДНК-полимераза присоединяет дезоксирибонуклеотидмонофосфат к 3’-ОН группе дезоксирибозы 5’ 5’ 3’ 3’ 5’ 5’ 3’
- 41. Репликация Лидирующая и отстающая нити ДНК 5’ 5’ 3’ 3’ 5’ 5’ 3’ 3’ Лидирующая нить
- 42. Репликация Фрагменты Оказаки Рэйдзи Оказаки (Reiji Okazaki) 1930—1975 Цунэко Оказаки (Tsuneko Okazaki) 1933 г.р. Ультрацентрифугирование ДНК
- 43. Репликация Фрагменты Оказаки Фрагменты Оказаки Вырезание праймеров и застройка бреши
- 45. Репликация Этапы репликации Расплетание ДНК Синтез праймера для начала считывания лидирующей цепи Синтез нуклеотидов + синтез
- 46. Репликация Этапы репликации Расплетание ДНК Топоизомераза (изомераза) Хеликаза (гидролаза) 5’ 5’ 3’ 3’ Белки SSB (single
- 47. Репликация Этапы репликации Синтез праймера для начала считывания лидирующей цепи Праймаза – ДНК-полимераза α (трансфераза) 5’
- 48. Репликация Этапы репликации Синтез нуклеотидов АДФ ГДФ ЦДФ Рибонуклеотид редуктаза (оксидоредуктаза) дАДФ дГДФ дЦДФ Нуклеозид-дифосфат киназа
- 49. Репликация Этапы репликации Синтез ДНК на лидирующей цепи ДНК-полимераза ε (трансфераза) 5’ 5’ 3’ 3’ ДНК-полимераза
- 50. Репликация Этапы репликации Синтез праймеров для отстающей цепи 5’ 5’ 3’ 3’ Праймаза – ДНК-полимераза α
- 51. Репликация Этапы репликации 5’ 5’ 3’ 3’ Синтез нуклеотидов + синтез фрагментов Оказаки ДНК-полимераза δ (трансфераза)
- 52. Репликация Этапы репликации Продвижение репликационной вилки (~1000 п.н.)
- 53. Репликация Этапы репликации Синтез праймеров для отстающей цепи Праймаза – ДНК-полимераза α (трансфераза)
- 54. Репликация Этапы репликации Синтез нуклеотидов + синтез фрагментов Оказаки ДНК-полимераза δ (трансфераза)
- 55. Репликация Этапы репликации Вырезание праймеров ДНК-полимераза δ вытесняет РНК праймер Эндонуклеаза (гидролаза) отрезает праймер Эндонуклеаза (гидролаза)
- 56. Репликация Этапы репликации Вырезание праймеров ДНК-лигаза (лигаза) застраивает одноцепочечный разрыв ДНК-лигаза
- 58. Репликация Скользящий зажим Белок β (E.coli) PCNA (эукариоты) Открытый зажим Закрытый зажим Закрытый зажим в зоне
- 59. Репликация Скользящий зажим Белок β и двухцепочечная ДНК Белок β PCNA
- 60. Репликация Скользящий зажим + PPi
- 61. Репликация Скользящий зажим ДНК-полимераза
- 62. Репликация Работа реплисомы TRENDS in Microbiology 2007 Vol.15 No.4 P.156-164
- 63. Репликация Выпетливание отстающей цепи
- 64. Репликация 3’→5’ экзонуклеазная активность ДНК-полимераз Одна ошибка на 105-106 н. Увеличение точности в 10-100 раз ДНК-полимеразы
- 66. Репликация Проблема концов хромосом эукариот Маргинотомия (1971) Лимит Хайфлика (1961) Нормальные (не опухолевые) клетки человека при
- 67. Репликация Теломеры τέλος — конец и μέρος — часть Концевые участки хромосом (TTAGGG)n 5’ 5’ 3’
- 68. Репликация Теломеры Укорачивание теломеров до n Укорачивание теломеров приводит к остановке деления (хотя маргинотомия не объясняет
- 69. Репликация Теломераза РНК-зависимая ДНК-полимераза (трансфераза) Добавляет фрагменты TTAGGG к 3’-концу хромосомы CAAUCCCAAUC TTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGG CAAUCCCAAUC TTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGG http://www.uic.edu
- 70. Репликация Теломераза Нобелевская премия 2009 г. за открытие механизмов защиты хромосом теломерами и фермента теломеразы
- 72. Клеточный цикл © Aaron Straight, Stanford University School of Medicine Интерфаза (I) Митоз (М)
- 73. Интерфаза Метафаза Профаза Прометафаза Анафаза Телофаза Клеточный цикл Митоз Alberts Bю et al. Molecular Biology of
- 74. Размер клетки Время I M I M Клеточный цикл Интерфаза Циклическое изменение размера делящейся клетки
- 75. Клеточный цикл Интерфаза Потребление клеткой дезоксинуклеотидов Время S фаза синтеза G1 1-я проме- жуточная фаза G2
- 76. Клеточный цикл G0 G1 G2 G0 S M Alberts B. et al. Molecular Biology of the
- 77. Клеточный цикл Исследование клеточного цикла http://www.cbp.pitt.edu/faculty/yong_wan/index.html
- 78. Клеточный цикл Исследование клеточного цикла Проточная цитофлуориметрия Спектр поглощения Спектр излучения Длина волны Интенсивность DAPI FITC
- 79. Количество ДНК в одной клетке Время S фаза синтеза G1 1-я проме- жуточная фаза G2 2-я
- 80. Клеточный цикл Исследование клеточного цикла Проточная цитофлуориметрия Схема прибора
- 81. Клеточный цикл Исследование клеточного цикла Проточная цитофлуориметрия Окрашивание клеточной ДНК йодистым пропидием PI Интенсивность флуоресценции Количество
- 82. Клеточный цикл Исследование клеточного цикла Интенсивность флуоресценции Количество клеток 2n 4n Окрашивание клеточной ДНК йодистым пропидием
- 83. Клеточный цикл Исследование клеточного цикла Проточная цитофлуориметрия Переход клеток в фазу G0 Типичные гистограммы окрашивания ДНК
- 85. Репликация Полимеразная цепная реакция (ПЦР) Матрица Симуляция работы хеликазы Симуляция работы праймазы Полимеризация ДНК …
- 86. Репликация Полимеразная цепная реакция (ПЦР) Симуляция работы хеликазы Плавление ДНК 96°С
- 87. Репликация Полимеразная цепная реакция (ПЦР) Симуляция работы праймазы Подбор и заказ праймеров Отжиг праймеров
- 88. Репликация Полимеразная цепная реакция (ПЦР) Полимеризация ДНК Thermus aquaticus Wikimedia Commons ДНК-полимераза Taq Tопт = 72°С
- 89. Репликация ПЦР 1-й цикл Плавление Отжиг Полимеризация 1 матрица (2 нити) + 1 полупродукт (2 нити)
- 90. Репликация ПЦР 2-й цикл Плавление Отжиг Полимеризация 1 матрица (2 нити) + 2 полупродукта (4 нити)
- 91. Репликация ПЦР 3-й цикл Плавление Отжиг Полимеризация 1 матрица (2 нити) + 3 полупродукта (6 нитей)
- 92. Репликация ПЦР Матриц - 1 Полупродуктов – n Продуктов – 2n-(n+1) Метод ПЦР позволяет определить наличие
- 94. Скачать презентацию

ФУНКЦИИ НУКЛЕИНОВЫХ КИСЛОТ
КАК ГЕНЕТИЧЕСКОГО МАТЕРИАЛА
Хранение, передача и реализация генетической информации
Хранение –
ФУНКЦИИ НУКЛЕИНОВЫХ КИСЛОТ
КАК ГЕНЕТИЧЕСКОГО МАТЕРИАЛА
Хранение, передача и реализация генетической информации
Хранение –
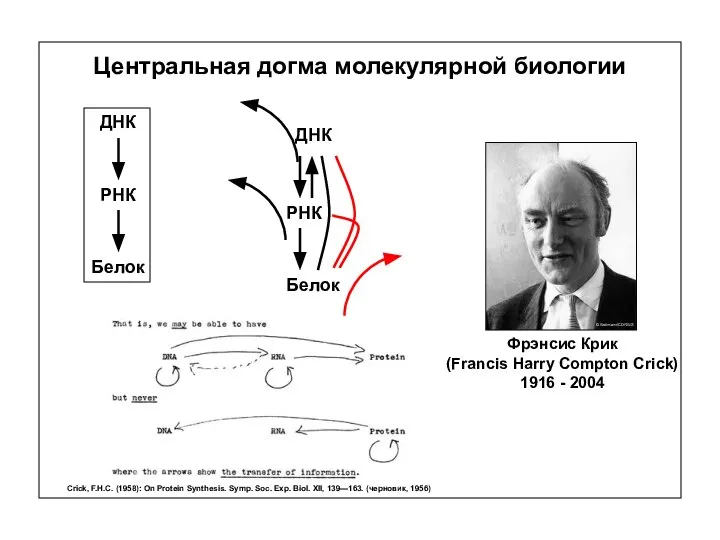
Центральная догма молекулярной биологии
Фрэнсис Крик
(Francis Harry Compton Crick)
1916 - 2004
ДНК
РНК
Белок
ДНК
РНК
Белок
Crick, F.H.C.
Центральная догма молекулярной биологии
Фрэнсис Крик
(Francis Harry Compton Crick)
1916 - 2004
ДНК
РНК
Белок
ДНК
РНК
Белок
Crick, F.H.C.
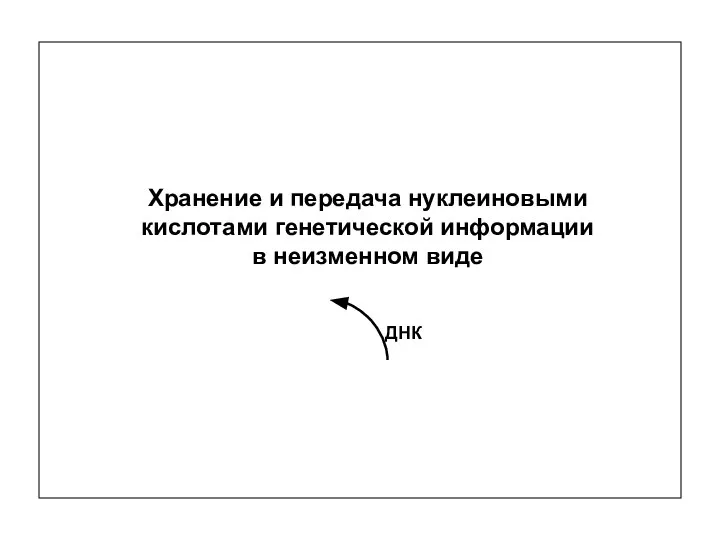
ДНК
Хранение и передача нуклеиновыми кислотами генетической информации
в неизменном виде
ДНК
Хранение и передача нуклеиновыми кислотами генетической информации
в неизменном виде
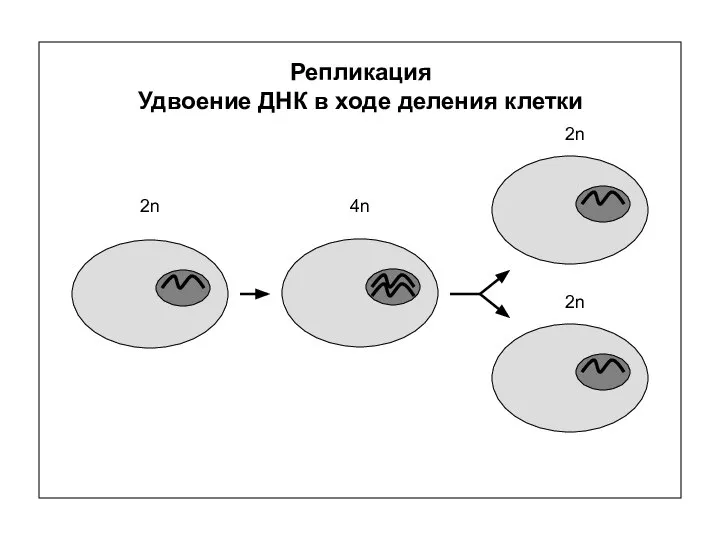
Репликация
Удвоение ДНК в ходе деления клетки
2n
4n
2n
2n
Репликация
Удвоение ДНК в ходе деления клетки
2n
4n
2n
2n
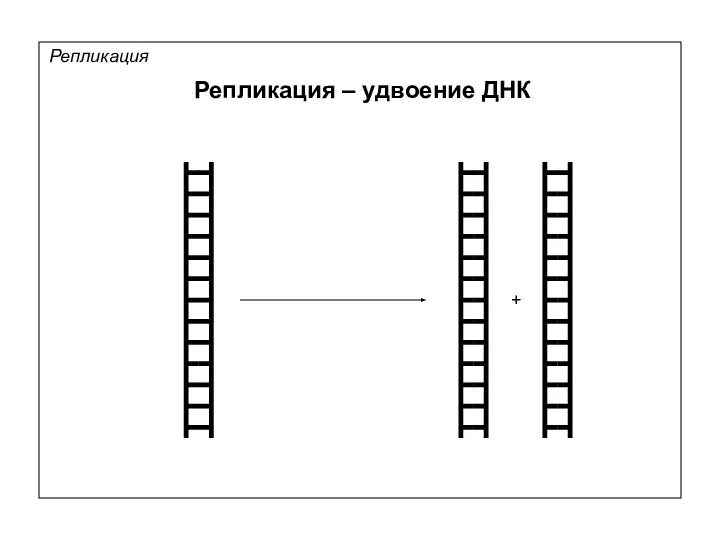
Репликация – удвоение ДНК
Репликация
+
Репликация – удвоение ДНК
Репликация
+
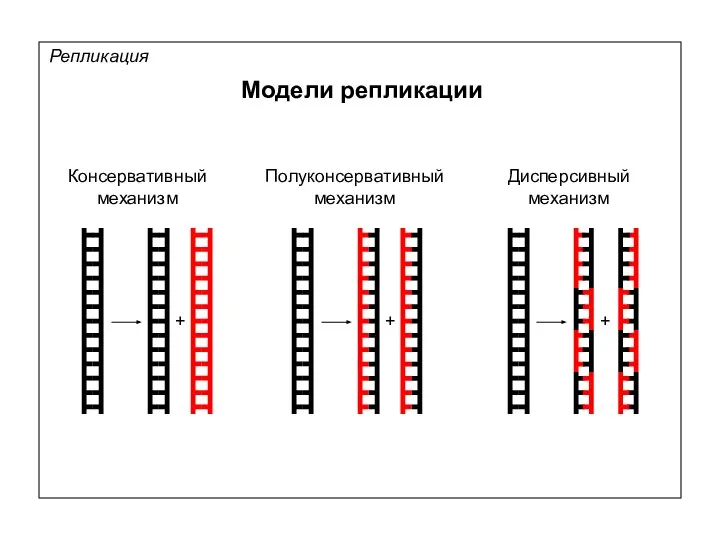
Модели репликации
Репликация
Консервативный
механизм
Полуконсервативный
механизм
Дисперсивный
механизм
+
+
+
Модели репликации
Репликация
Консервативный
механизм
Полуконсервативный
механизм
Дисперсивный
механизм
+
+
+

Полуконсервативный механизм репликации
Репликация
Мэтью Стэнли Мезенсон
Matthew Stanley Meselson
1930 г.р.
Франклин Уильям Сталь
Franklin William
Полуконсервативный механизм репликации
Репликация
Мэтью Стэнли Мезенсон
Matthew Stanley Meselson
1930 г.р.
Франклин Уильям Сталь
Franklin William

Эксперимент Мезельсона-Сталя (PNAS.-1958.-44:671-682)
Репликация
14N - 99,635 % и 15N - 0,365 %
15NH4Cl
E.coli
E.coli
14NH4Cl
ДНК
cодержит
Эксперимент Мезельсона-Сталя (PNAS.-1958.-44:671-682)
Репликация
14N - 99,635 % и 15N - 0,365 %
15NH4Cl
E.coli
E.coli
14NH4Cl
ДНК
cодержит
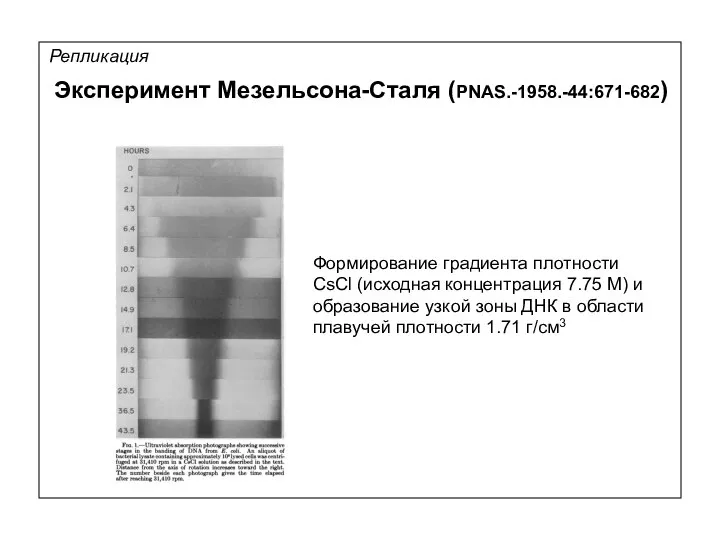
Репликация
Формирование градиента плотности
CsCl (исходная концентрация 7.75 М) и
образование узкой зоны ДНК
Репликация
Формирование градиента плотности
CsCl (исходная концентрация 7.75 М) и
образование узкой зоны ДНК
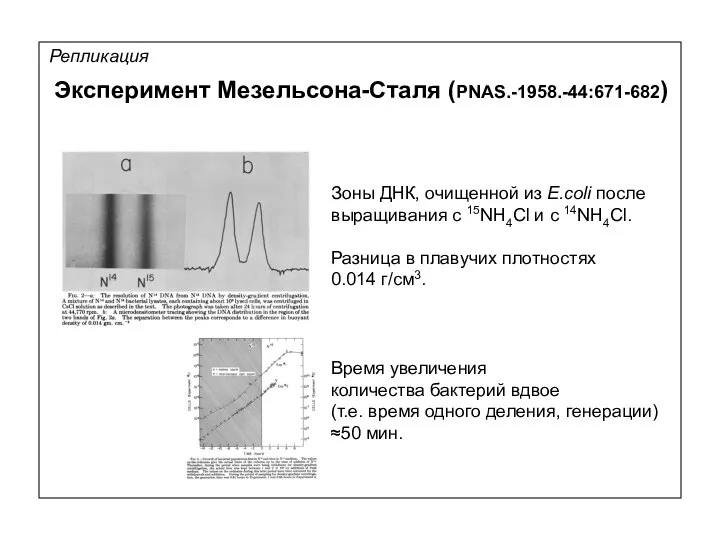
Репликация
Зоны ДНК, очищенной из E.coli после
выращивания с 15NH4Cl и c 14NH4Cl.
Разница
Репликация
Зоны ДНК, очищенной из E.coli после
выращивания с 15NH4Cl и c 14NH4Cl.
Разница

Репликация
ДНК из E.coli после выращивания с 15NH4Cl
ДНК из E.coli после
Репликация
ДНК из E.coli после выращивания с 15NH4Cl
ДНК из E.coli после
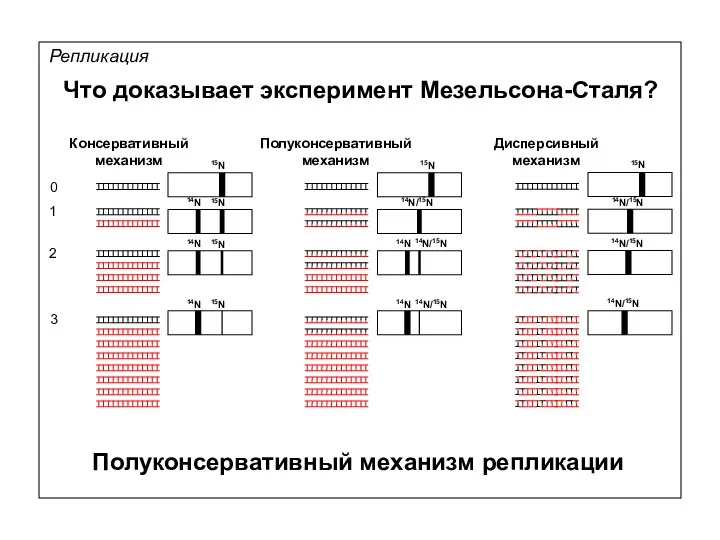
Репликация
Что доказывает эксперимент Мезельсона-Сталя?
0
1
2
3
Дисперсивный
механизм
15N
15N
14N
15N
14N
15N
14N
15N
14N/15N
14N
14N
14N/15N
14N/15N
15N
14N/15N
14N/15N
14N/15N
Полуконсервативный механизм репликации
Репликация
Что доказывает эксперимент Мезельсона-Сталя?
0
1
2
3
Дисперсивный
механизм
15N
15N
14N
15N
14N
15N
14N
15N
14N/15N
14N
14N
14N/15N
14N/15N
15N
14N/15N
14N/15N
14N/15N
Полуконсервативный механизм репликации
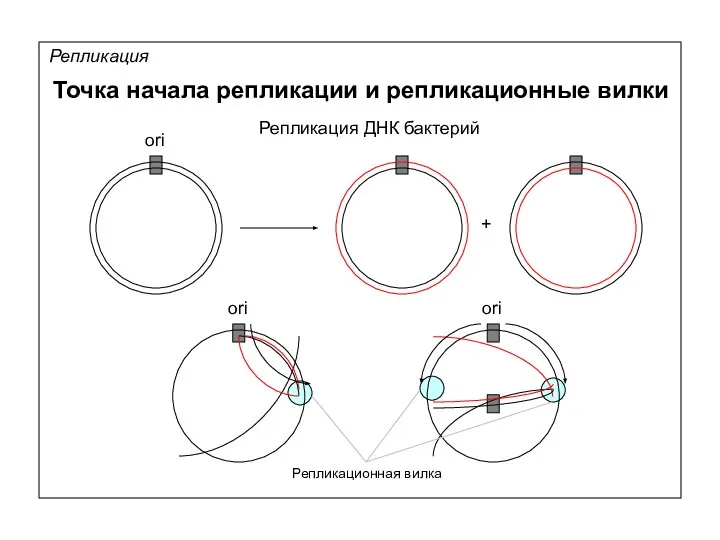
Репликация
Точка начала репликации и репликационные вилки
ori
Репликация ДНК бактерий
+
Репликационная вилка
Репликация
Точка начала репликации и репликационные вилки
ori
Репликация ДНК бактерий
+
Репликационная вилка
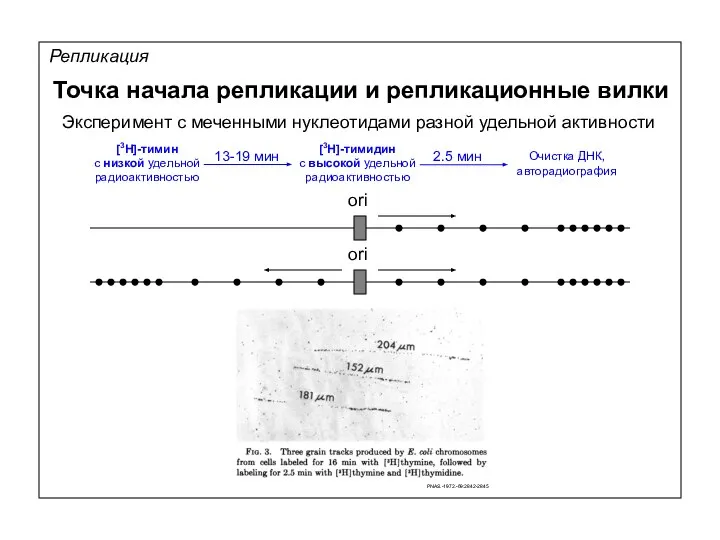
13-19 мин
Репликация
Точка начала репликации и репликационные вилки
ori
ori
Эксперимент с меченными нуклеотидами разной
13-19 мин
Репликация
Точка начала репликации и репликационные вилки
ori
ori
Эксперимент с меченными нуклеотидами разной
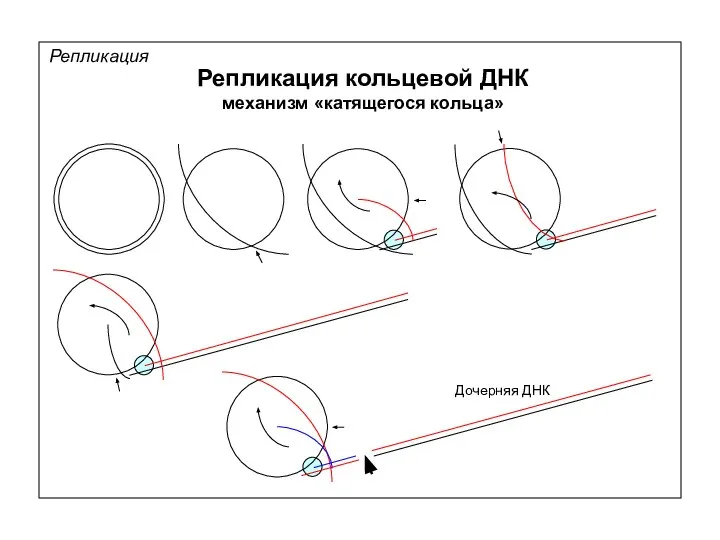
Репликация
Репликация кольцевой ДНК
механизм «катящегося кольца»
Дочерняя ДНК
Репликация
Репликация кольцевой ДНК
механизм «катящегося кольца»
Дочерняя ДНК
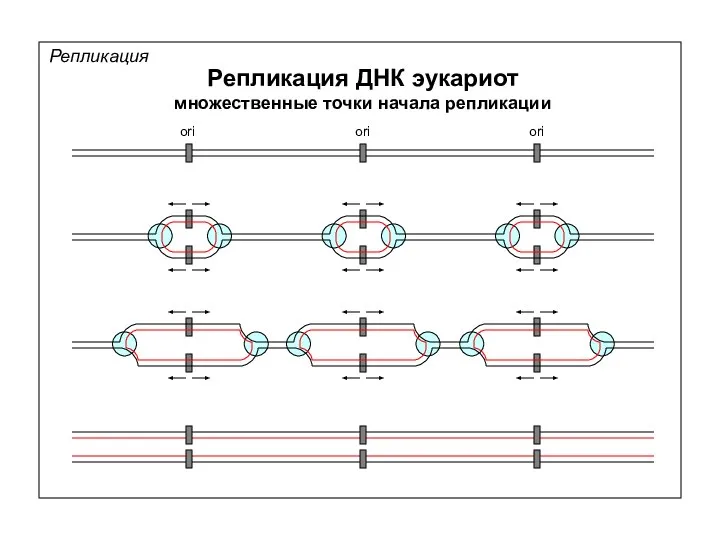
Репликация
Репликация ДНК эукариот
множественные точки начала репликации
ori
ori
ori
Репликация
Репликация ДНК эукариот
множественные точки начала репликации
ori
ori
ori
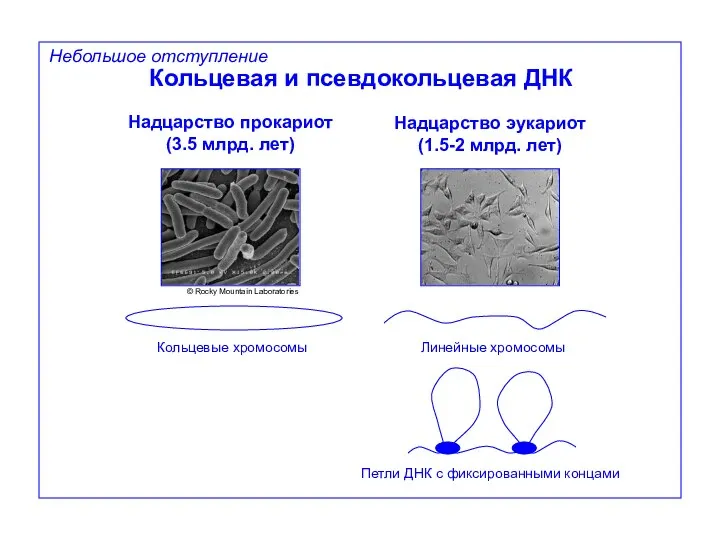
Небольшое отступление
Кольцевая и псевдокольцевая ДНК
Надцарство прокариот
(3.5 млрд. лет)
Надцарство эукариот
(1.5-2 млрд. лет)
©
Небольшое отступление
Кольцевая и псевдокольцевая ДНК
Надцарство прокариот
(3.5 млрд. лет)
Надцарство эукариот
(1.5-2 млрд. лет)
©


1. Оксидоредуктазы
2. Трансферазы
3. Гидролазы
4. Лиазы
5. Изомеразы
6. Лигазы
Небольшое отступление
A(окисл) + B(восст) →
1. Оксидоредуктазы
2. Трансферазы
3. Гидролазы
4. Лиазы
5. Изомеразы
6. Лигазы
Небольшое отступление
A(окисл) + B(восст) →
![Репликация ДНК-полимераза (трансфераза) α-[32P]-дезоксирибонуклеотиды α-[32P]-дАТФ α-[32P]-дГТФ α-[32P]-дЦТФ α-[32P]-дТТФ + экстракт E.coli Очистка ДНК Измерение радиоактивности ДНК](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1395811/slide-20.jpg)
Репликация
ДНК-полимераза
(трансфераза)
α-[32P]-дезоксирибонуклеотиды
α-[32P]-дАТФ
α-[32P]-дГТФ
α-[32P]-дЦТФ
α-[32P]-дТТФ
+ экстракт E.coli
Очистка ДНК
Измерение
радиоактивности
ДНК
Репликация
ДНК-полимераза
(трансфераза)
α-[32P]-дезоксирибонуклеотиды
α-[32P]-дАТФ
α-[32P]-дГТФ
α-[32P]-дЦТФ
α-[32P]-дТТФ
+ экстракт E.coli
Очистка ДНК
Измерение
радиоактивности
ДНК
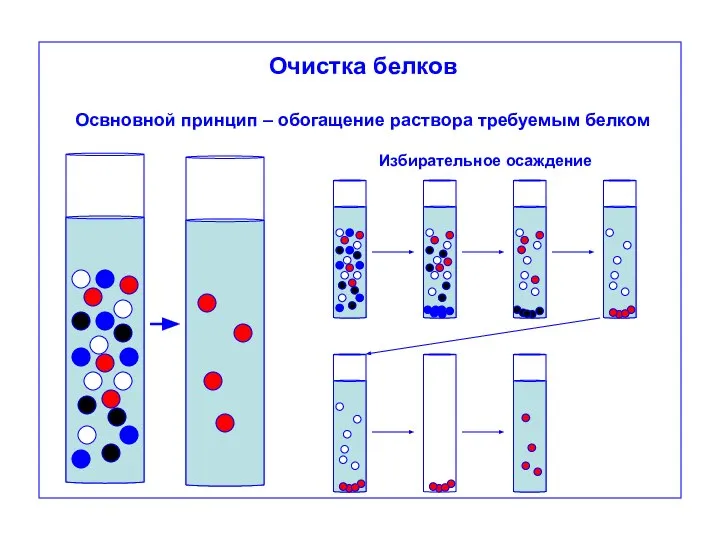
Очистка белков
Освновной принцип – обогащение раствора требуемым белком
Избирательное осаждение
Очистка белков
Освновной принцип – обогащение раствора требуемым белком
Избирательное осаждение
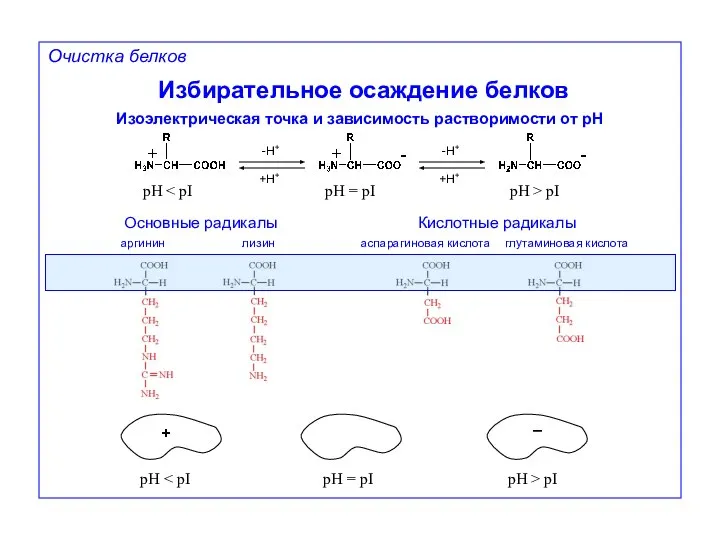
Очистка белков
Избирательное осаждение белков
Изоэлектрическая точка и зависимость растворимости от pH
-H+
+H+
-H+
+H+
pH <
Очистка белков
Избирательное осаждение белков
Изоэлектрическая точка и зависимость растворимости от pH
-H+
+H+
-H+
+H+
pH <
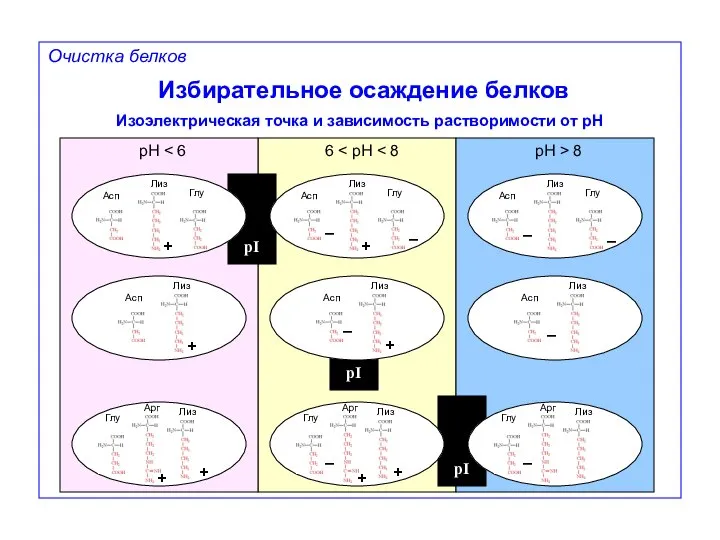
Избирательное осаждение белков
Изоэлектрическая точка и зависимость растворимости от pH
pH < 6
6
Избирательное осаждение белков
Изоэлектрическая точка и зависимость растворимости от pH
pH < 6
6
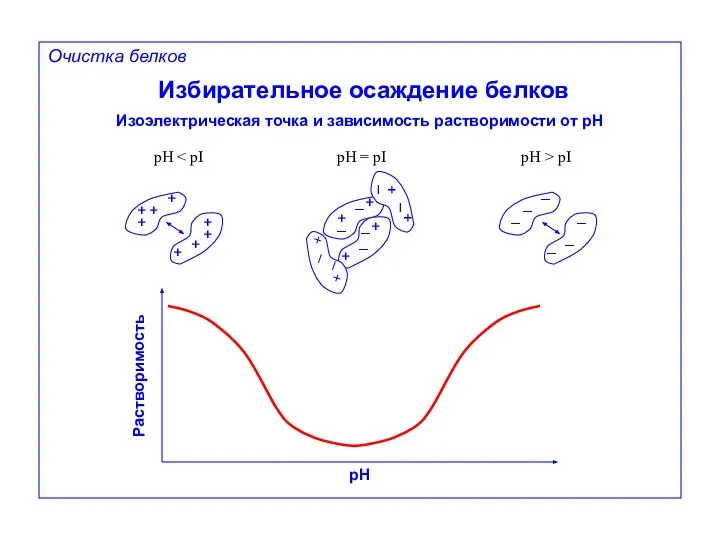
Избирательное осаждение белков
Изоэлектрическая точка и зависимость растворимости от pH
+
+
+
+
+
+
–
–
–
–
–
–
+
+
pH < pI
pH
Избирательное осаждение белков
Изоэлектрическая точка и зависимость растворимости от pH
+
+
+
+
+
+
–
–
–
–
–
–
+
+
pH < pI
pH
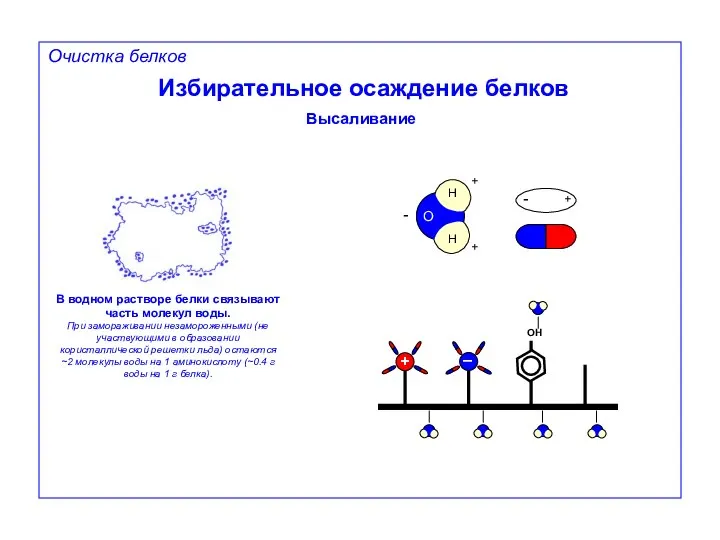
Избирательное осаждение белков
Высаливание
В водном растворе белки связывают часть молекул воды.
При замораживании
Избирательное осаждение белков
Высаливание
В водном растворе белки связывают часть молекул воды.
При замораживании
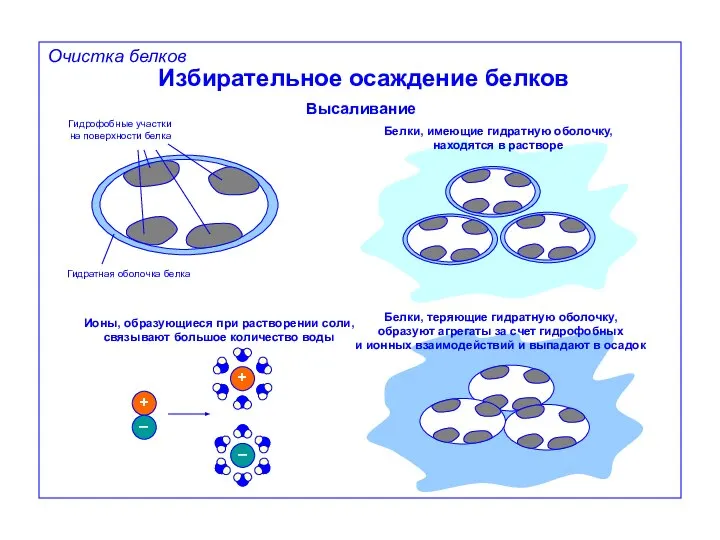
Избирательное осаждение белков
Высаливание
Гидрофобные участки
на поверхности белка
Гидратная оболочка белка
Белки, имеющие гидратную оболочку,
находятся
Избирательное осаждение белков
Высаливание
Гидрофобные участки
на поверхности белка
Гидратная оболочка белка
Белки, имеющие гидратную оболочку,
находятся
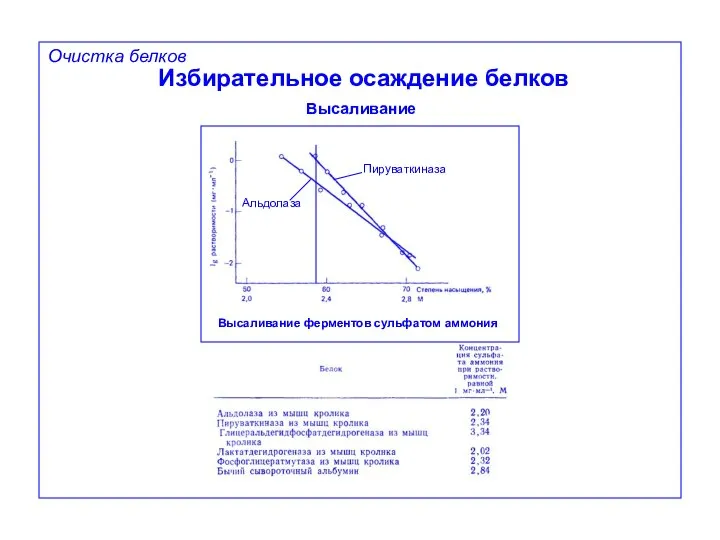
Избирательное осаждение белков
Высаливание
Очистка белков
Избирательное осаждение белков
Высаливание
Очистка белков

Избирательное осаждение белков
Денатурация
(удаление из смеси ненужных белков)
pH (сильный сдвиг)
Температура
Органические растворители
Денатурация
(потеря третичной
Избирательное осаждение белков
Денатурация
(удаление из смеси ненужных белков)
pH (сильный сдвиг)
Температура
Органические растворители
Денатурация
(потеря третичной
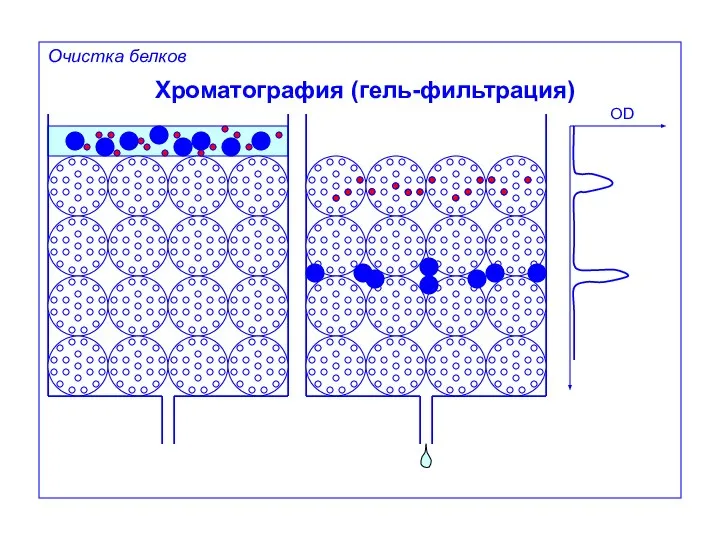
Хроматография (гель-фильтрация)
OD
Очистка белков
Хроматография (гель-фильтрация)
OD
Очистка белков
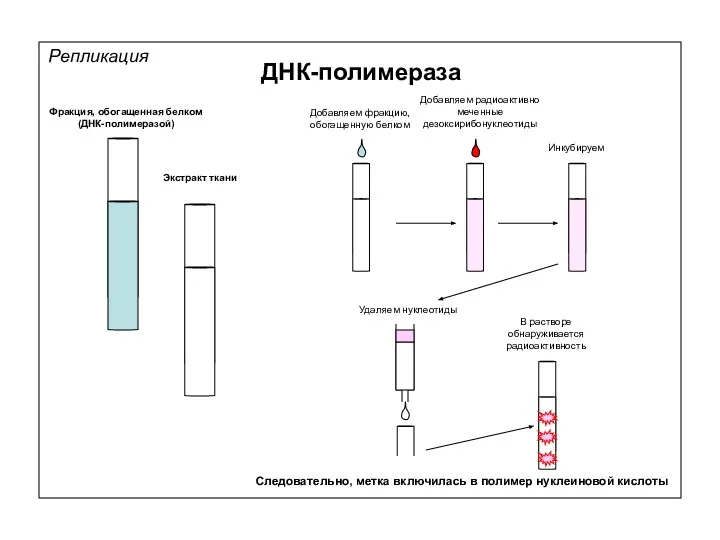
Репликация
ДНК-полимераза
Фракция, обогащенная белком
(ДНК-полимеразой)
Экстракт ткани
Добавляем фракцию, обогащенную белком
Добавляем радиоактивно меченные дезоксирибонуклеотиды
Инкубируем
Удаляем нуклеотиды
В
Репликация
ДНК-полимераза
Фракция, обогащенная белком
(ДНК-полимеразой)
Экстракт ткани
Добавляем фракцию, обогащенную белком
Добавляем радиоактивно меченные дезоксирибонуклеотиды
Инкубируем
Удаляем нуклеотиды
В
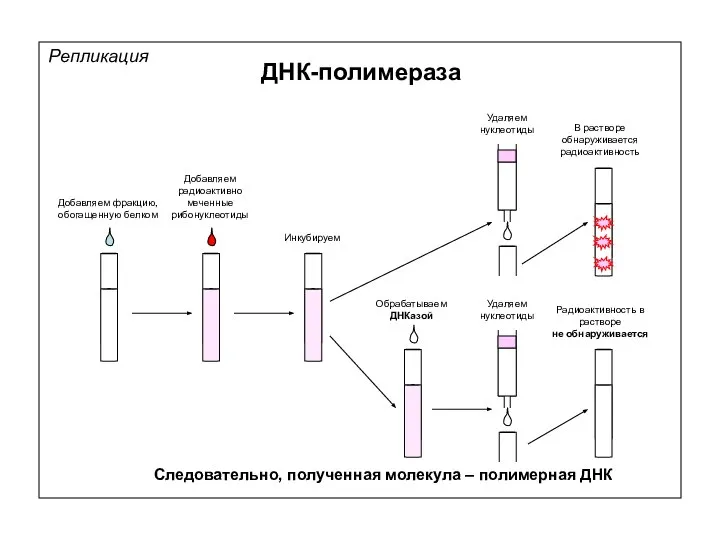
Удаляем
нуклеотиды
Удаляем
нуклеотиды
Добавляем фракцию, обогащенную белком
Добавляем радиоактивно меченные рибонуклеотиды
Инкубируем
Радиоактивность в растворе
не обнаруживается
Следовательно, полученная
Удаляем
нуклеотиды
Удаляем
нуклеотиды
Добавляем фракцию, обогащенную белком
Добавляем радиоактивно меченные рибонуклеотиды
Инкубируем
Радиоактивность в растворе
не обнаруживается
Следовательно, полученная
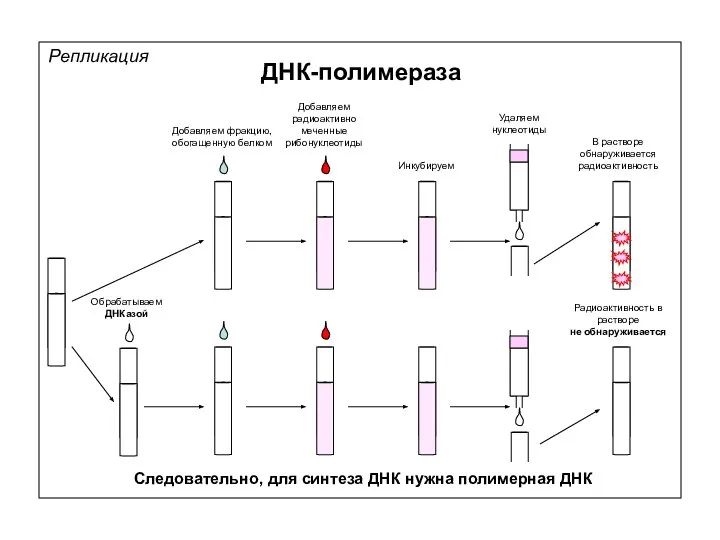
Добавляем фракцию, обогащенную белком
Добавляем радиоактивно меченные рибонуклеотиды
Инкубируем
Следовательно, для синтеза ДНК нужна
Добавляем фракцию, обогащенную белком
Добавляем радиоактивно меченные рибонуклеотиды
Инкубируем
Следовательно, для синтеза ДНК нужна
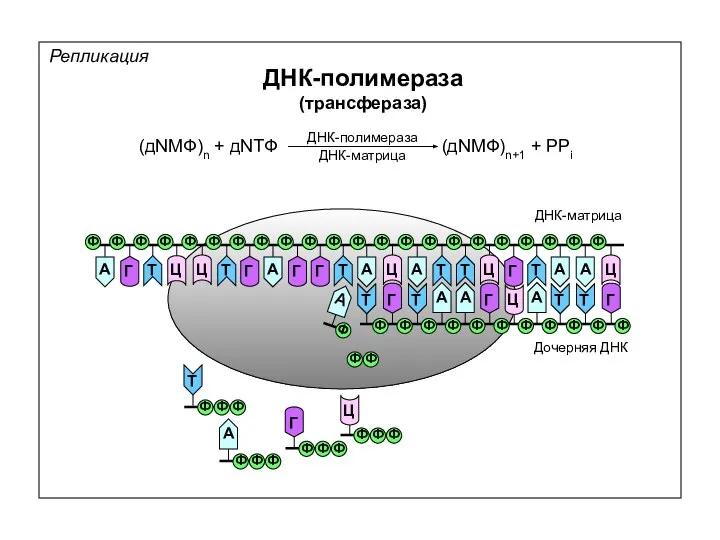
Репликация
ДНК-полимераза
(трансфераза)
(дNМФ)n + дNТФ
(дNМФ)n+1 + PPi
ДНК-полимераза
ДНК-матрица
ДНК-матрица
Дочерняя ДНК
Репликация
ДНК-полимераза
(трансфераза)
(дNМФ)n + дNТФ
(дNМФ)n+1 + PPi
ДНК-полимераза
ДНК-матрица
ДНК-матрица
Дочерняя ДНК


ДНК-полимераза требует наличия матрицы (материнской ДНК).
ДНК-полимераза присоединяет дезоксирибонуклеозидмонофосфат в соответствии с
ДНК-полимераза требует наличия матрицы (материнской ДНК).
ДНК-полимераза присоединяет дезоксирибонуклеозидмонофосфат в соответствии с
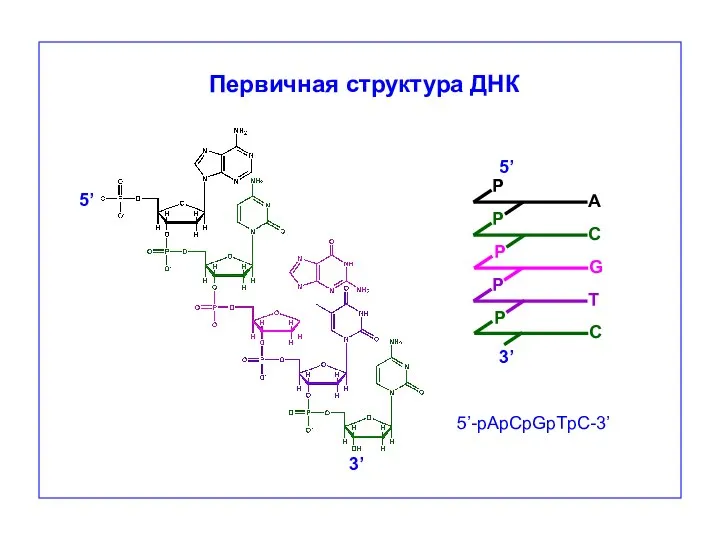
P
G
5’
3’
5’-pApCpGpTpC-3’
P
P
A
C
P
C
P
T
3’
5’
Первичная структура ДНК
P
G
5’
3’
5’-pApCpGpTpC-3’
P
P
A
C
P
C
P
T
3’
5’
Первичная структура ДНК
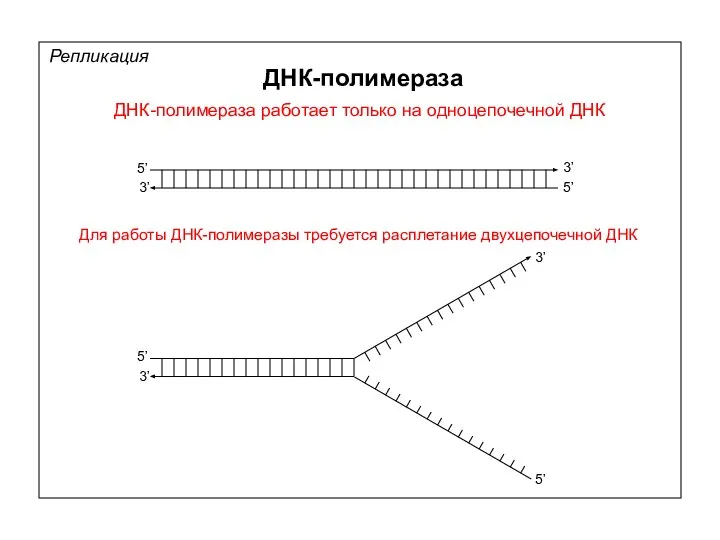
ДНК-полимераза работает только на одноцепочечной ДНК
Репликация
ДНК-полимераза
5’
5’
3’
3’
Для работы ДНК-полимеразы требуется расплетание двухцепочечной
ДНК-полимераза работает только на одноцепочечной ДНК
Репликация
ДНК-полимераза
5’
5’
3’
3’
Для работы ДНК-полимеразы требуется расплетание двухцепочечной

ДНК-полимераза не может начать синтез ДНК с первого нуклеотида, она присоединяет
ДНК-полимераза не может начать синтез ДНК с первого нуклеотида, она присоединяет
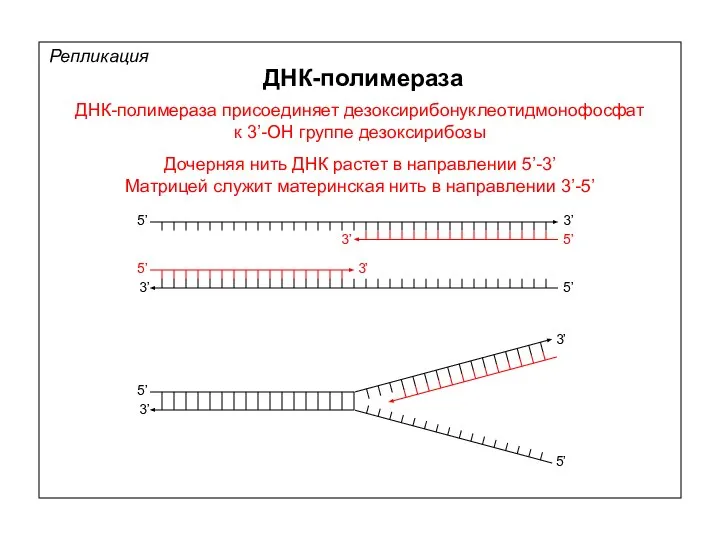
Репликация
ДНК-полимераза
ДНК-полимераза присоединяет дезоксирибонуклеотидмонофосфат
к 3’-ОН группе дезоксирибозы
5’
5’
3’
3’
5’
5’
3’
3’
Дочерняя нить ДНК растет в направлении
Репликация
ДНК-полимераза
ДНК-полимераза присоединяет дезоксирибонуклеотидмонофосфат
к 3’-ОН группе дезоксирибозы
5’
5’
3’
3’
5’
5’
3’
3’
Дочерняя нить ДНК растет в направлении
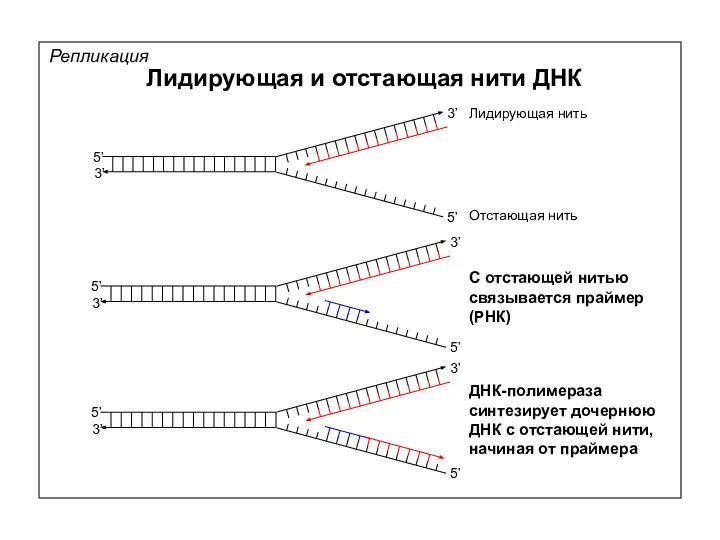
Репликация
Лидирующая и отстающая нити ДНК
5’
5’
3’
3’
5’
5’
3’
3’
Лидирующая нить
Отстающая нить
С отстающей нитью связывается праймер
Репликация
Лидирующая и отстающая нити ДНК
5’
5’
3’
3’
5’
5’
3’
3’
Лидирующая нить
Отстающая нить
С отстающей нитью связывается праймер
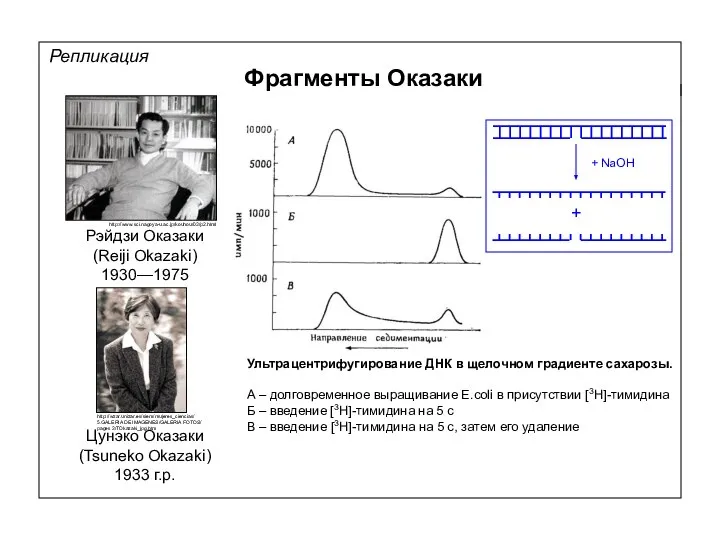
Репликация
Фрагменты Оказаки
Рэйдзи Оказаки
(Reiji Okazaki)
1930—1975
Цунэко Оказаки
(Tsuneko Okazaki)
1933 г.р.
Ультрацентрифугирование ДНК в щелочном градиенте
Репликация
Фрагменты Оказаки
Рэйдзи Оказаки
(Reiji Okazaki)
1930—1975
Цунэко Оказаки
(Tsuneko Okazaki)
1933 г.р.
Ультрацентрифугирование ДНК в щелочном градиенте
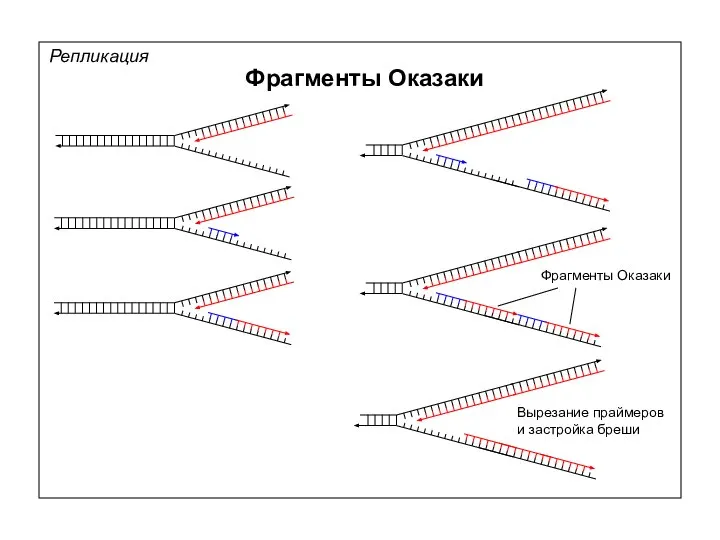
Репликация
Фрагменты Оказаки
Фрагменты Оказаки
Вырезание праймеров
и застройка бреши
Репликация
Фрагменты Оказаки
Фрагменты Оказаки
Вырезание праймеров
и застройка бреши

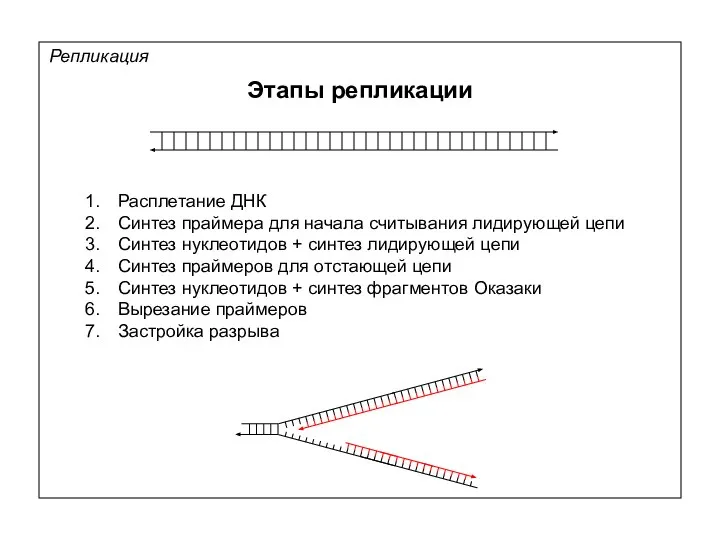
Репликация
Этапы репликации
Расплетание ДНК
Синтез праймера для начала считывания лидирующей цепи
Синтез нуклеотидов +
Репликация
Этапы репликации
Расплетание ДНК
Синтез праймера для начала считывания лидирующей цепи
Синтез нуклеотидов +
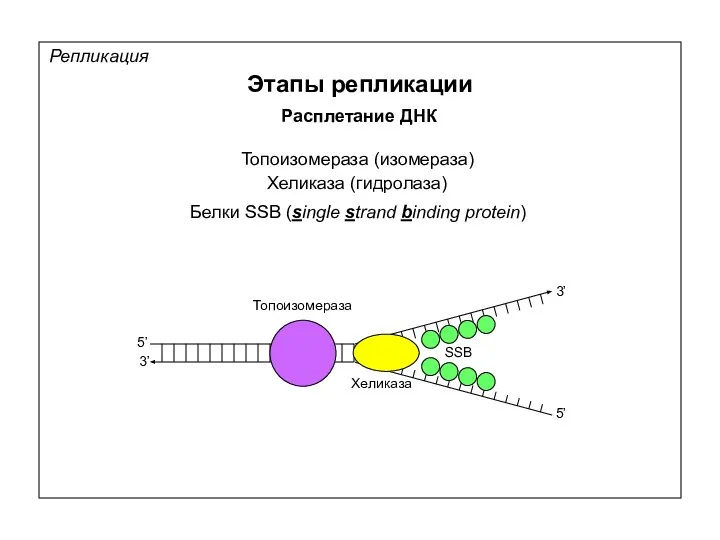
Репликация
Этапы репликации
Расплетание ДНК
Топоизомераза (изомераза)
Хеликаза (гидролаза)
5’
5’
3’
3’
Белки SSB (single strand binding protein)
Топоизомераза
Хеликаза
SSB
Репликация
Этапы репликации
Расплетание ДНК
Топоизомераза (изомераза)
Хеликаза (гидролаза)
5’
5’
3’
3’
Белки SSB (single strand binding protein)
Топоизомераза
Хеликаза
SSB
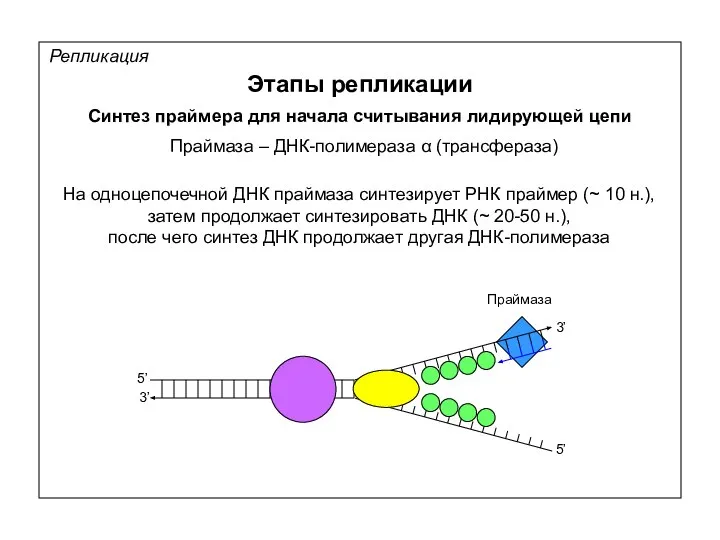
Репликация
Этапы репликации
Синтез праймера для начала считывания лидирующей цепи
Праймаза – ДНК-полимераза α
Репликация
Этапы репликации
Синтез праймера для начала считывания лидирующей цепи
Праймаза – ДНК-полимераза α
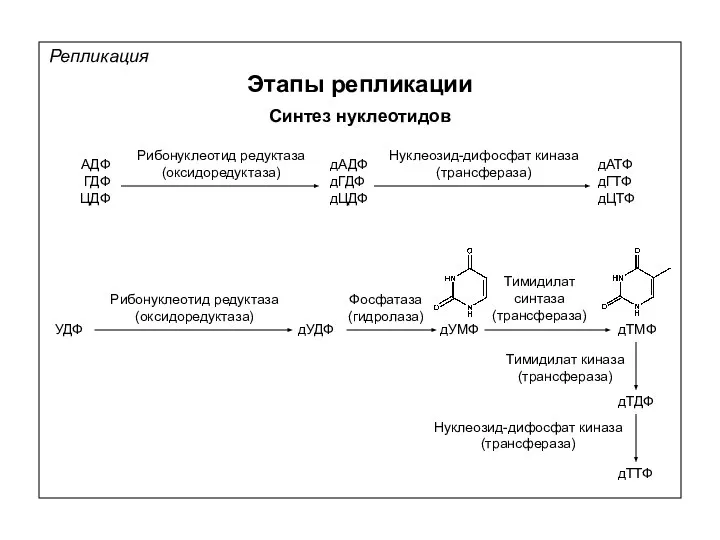
Репликация
Этапы репликации
Синтез нуклеотидов
АДФ
ГДФ
ЦДФ
Рибонуклеотид редуктаза
(оксидоредуктаза)
дАДФ
дГДФ
дЦДФ
Нуклеозид-дифосфат киназа
(трансфераза)
дАТФ
дГТФ
дЦТФ
УДФ
Рибонуклеотид редуктаза
(оксидоредуктаза)
дУДФ
Фосфатаза
(гидролаза)
дУМФ
Тимидилат
синтаза
(трансфераза)
дТМФ
дТДФ
дТТФ
Тимидилат киназа
(трансфераза)
Нуклеозид-дифосфат киназа
(трансфераза)
Репликация
Этапы репликации
Синтез нуклеотидов
АДФ
ГДФ
ЦДФ
Рибонуклеотид редуктаза
(оксидоредуктаза)
дАДФ
дГДФ
дЦДФ
Нуклеозид-дифосфат киназа
(трансфераза)
дАТФ
дГТФ
дЦТФ
УДФ
Рибонуклеотид редуктаза
(оксидоредуктаза)
дУДФ
Фосфатаза
(гидролаза)
дУМФ
Тимидилат
синтаза
(трансфераза)
дТМФ
дТДФ
дТТФ
Тимидилат киназа
(трансфераза)
Нуклеозид-дифосфат киназа
(трансфераза)
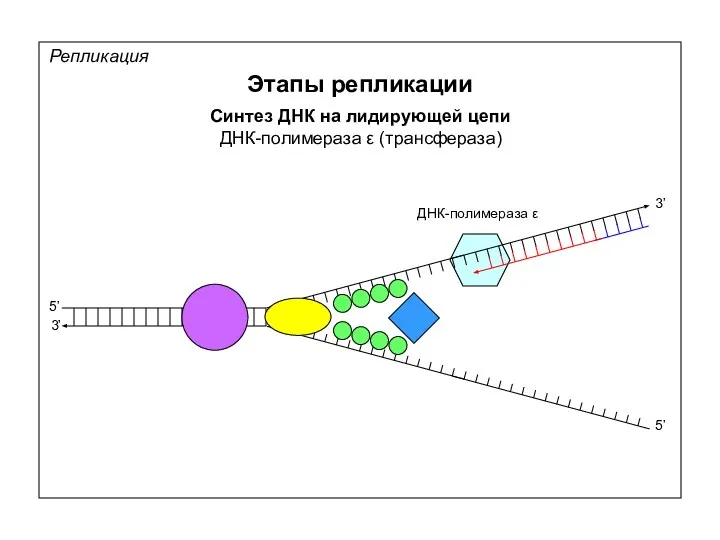
Репликация
Этапы репликации
Синтез ДНК на лидирующей цепи
ДНК-полимераза ε (трансфераза)
5’
5’
3’
3’
ДНК-полимераза ε
Репликация
Этапы репликации
Синтез ДНК на лидирующей цепи
ДНК-полимераза ε (трансфераза)
5’
5’
3’
3’
ДНК-полимераза ε
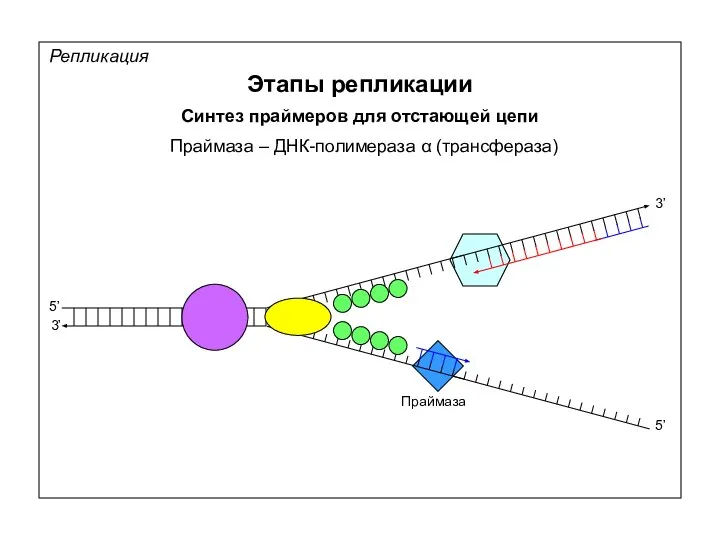
Репликация
Этапы репликации
Синтез праймеров для отстающей цепи
5’
5’
3’
3’
Праймаза – ДНК-полимераза α (трансфераза)
Праймаза
Репликация
Этапы репликации
Синтез праймеров для отстающей цепи
5’
5’
3’
3’
Праймаза – ДНК-полимераза α (трансфераза)
Праймаза
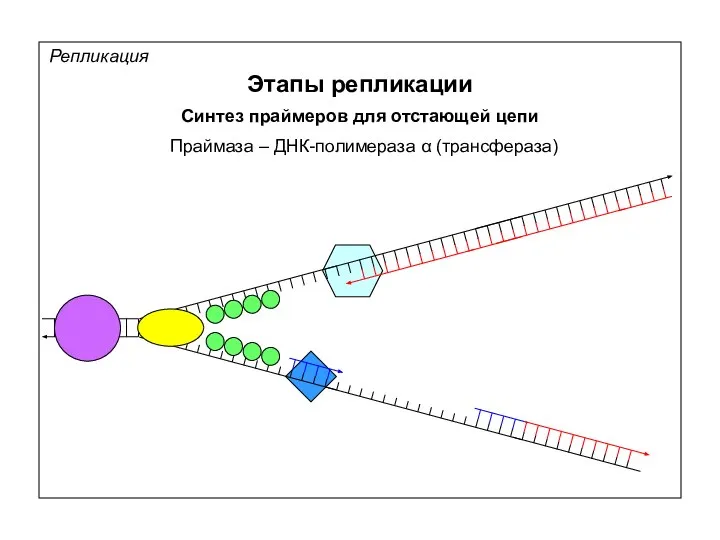
Репликация
Этапы репликации
5’
5’
3’
3’
Синтез нуклеотидов + синтез фрагментов Оказаки
ДНК-полимераза δ (трансфераза)
ДНК-полимераза δ
Репликация
Этапы репликации
5’
5’
3’
3’
Синтез нуклеотидов + синтез фрагментов Оказаки
ДНК-полимераза δ (трансфераза)
ДНК-полимераза δ
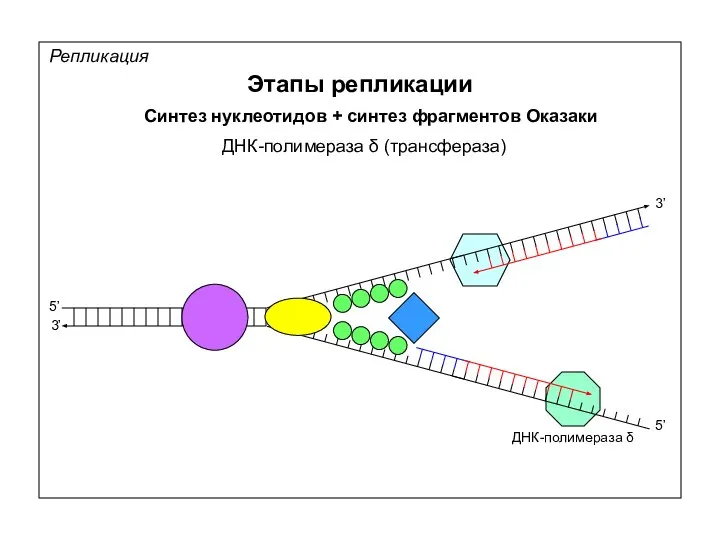
Репликация
Этапы репликации
Продвижение репликационной вилки (~1000 п.н.)
Репликация
Этапы репликации
Продвижение репликационной вилки (~1000 п.н.)
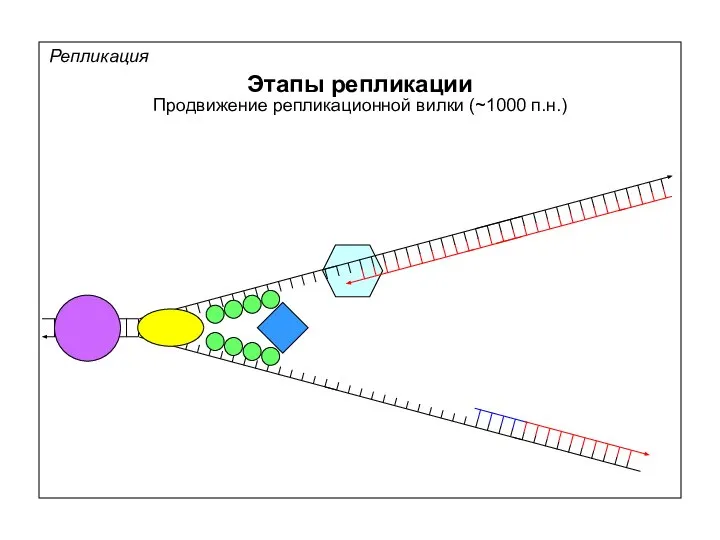
Репликация
Этапы репликации
Синтез праймеров для отстающей цепи
Праймаза – ДНК-полимераза α (трансфераза)
Репликация
Этапы репликации
Синтез праймеров для отстающей цепи
Праймаза – ДНК-полимераза α (трансфераза)
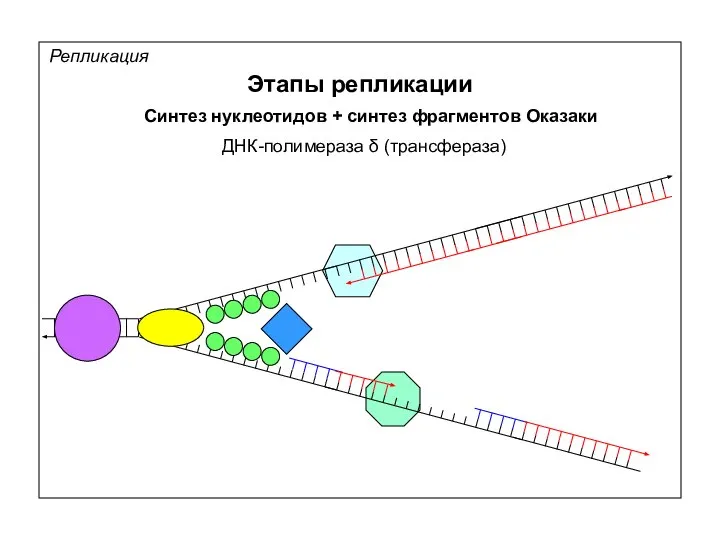
Репликация
Этапы репликации
Синтез нуклеотидов + синтез фрагментов Оказаки
ДНК-полимераза δ (трансфераза)
Репликация
Этапы репликации
Синтез нуклеотидов + синтез фрагментов Оказаки
ДНК-полимераза δ (трансфераза)
Репликация
Этапы репликации
Вырезание праймеров
ДНК-полимераза δ вытесняет РНК праймер
Эндонуклеаза (гидролаза)
отрезает праймер
Эндонуклеаза
(гидролаза)
Однонитевой
разрыв
Репликация
Этапы репликации
Вырезание праймеров
ДНК-полимераза δ вытесняет РНК праймер
Эндонуклеаза (гидролаза)
отрезает праймер
Эндонуклеаза
(гидролаза)
Однонитевой
разрыв
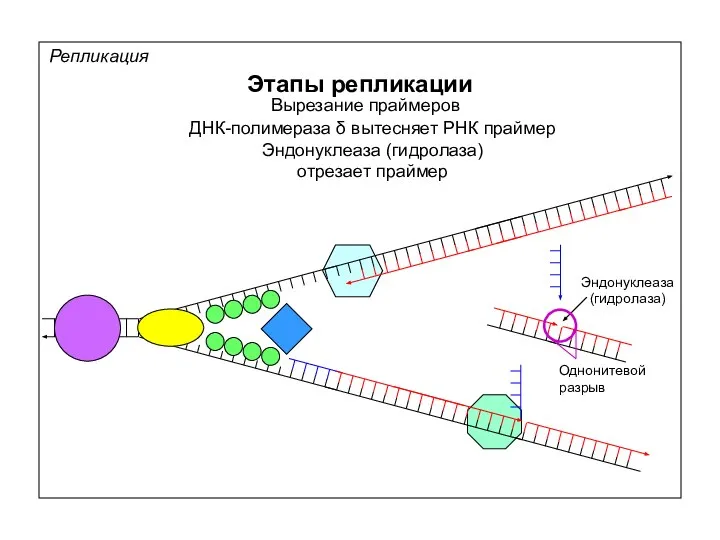
Репликация
Этапы репликации
Вырезание праймеров
ДНК-лигаза (лигаза) застраивает одноцепочечный разрыв
ДНК-лигаза
Репликация
Этапы репликации
Вырезание праймеров
ДНК-лигаза (лигаза) застраивает одноцепочечный разрыв
ДНК-лигаза

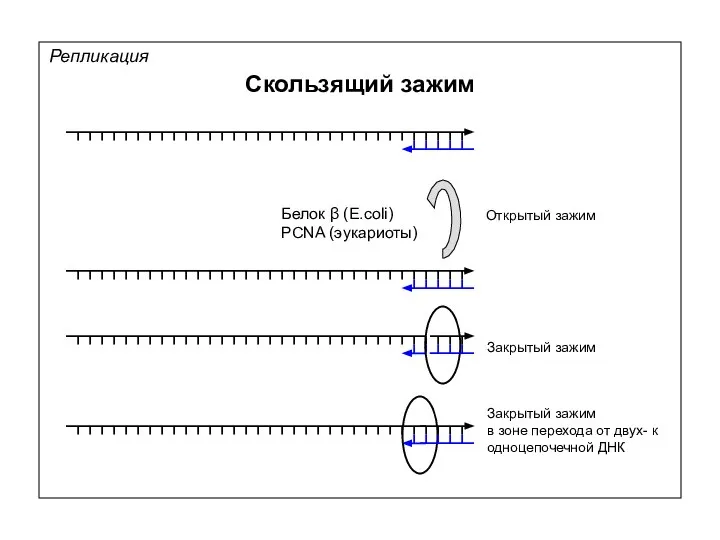
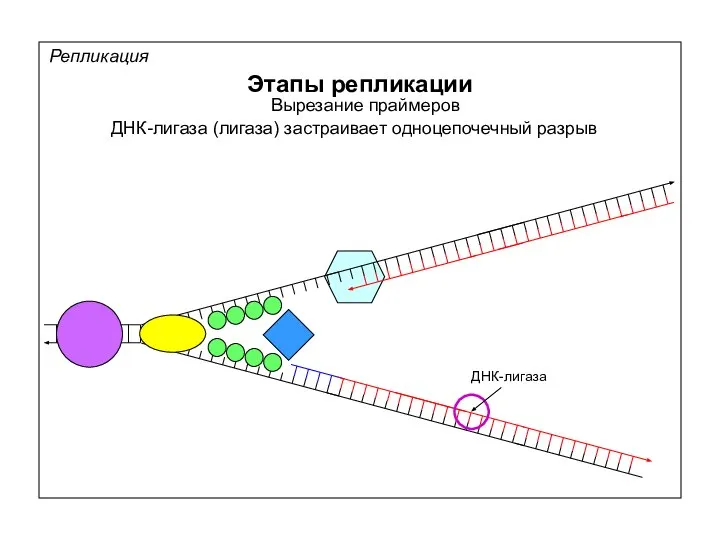
Репликация
Скользящий зажим
Белок β (E.coli)
PCNA (эукариоты)
Открытый зажим
Закрытый зажим
Закрытый зажим
в зоне перехода от
Репликация
Скользящий зажим
Белок β (E.coli)
PCNA (эукариоты)
Открытый зажим
Закрытый зажим
Закрытый зажим
в зоне перехода от
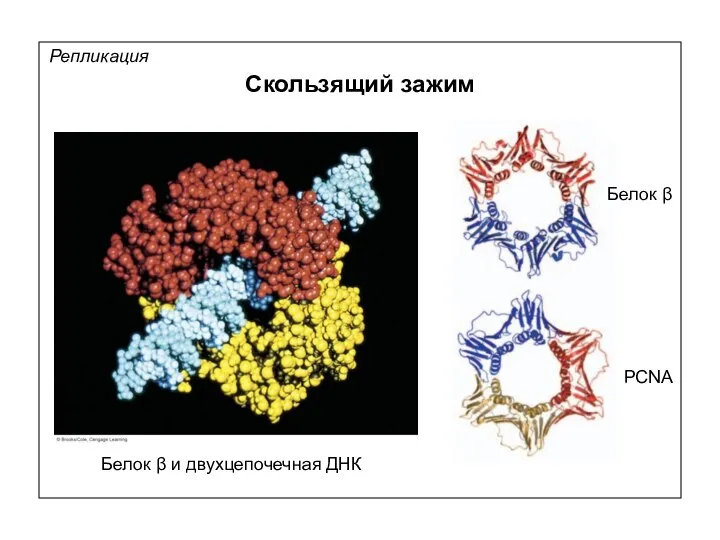
Репликация
Скользящий зажим
Белок β и двухцепочечная ДНК
Белок β
PCNA
Репликация
Скользящий зажим
Белок β и двухцепочечная ДНК
Белок β
PCNA
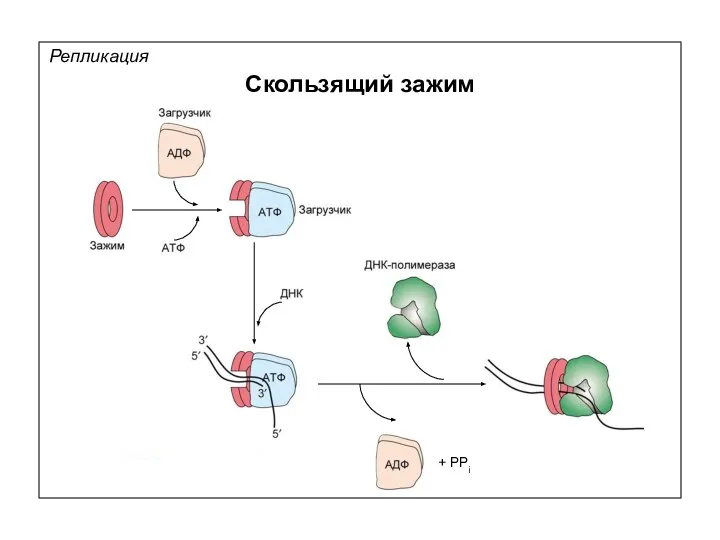
Репликация
Скользящий зажим
+ PPi
Репликация
Скользящий зажим
+ PPi
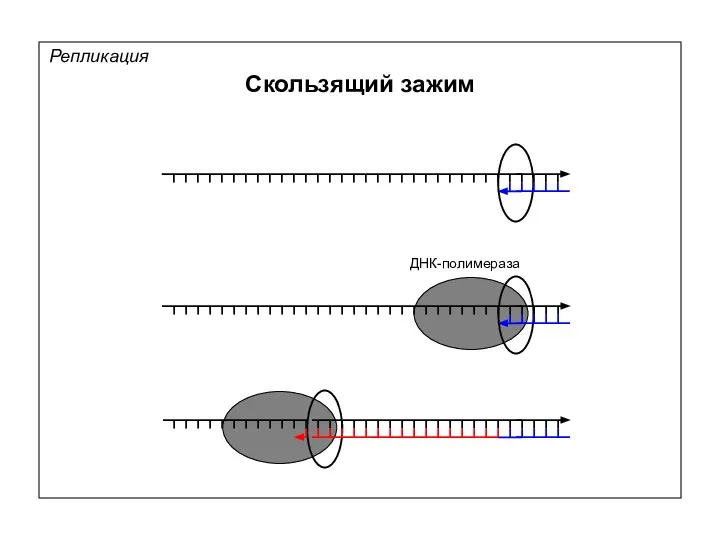
Репликация
Скользящий зажим
ДНК-полимераза
Репликация
Скользящий зажим
ДНК-полимераза
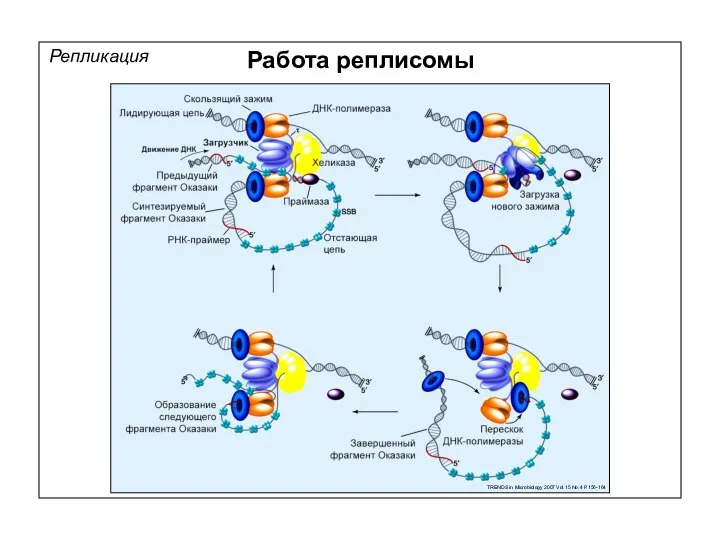
Репликация
Работа реплисомы
TRENDS in Microbiology 2007 Vol.15 No.4 P.156-164
Репликация
Работа реплисомы
TRENDS in Microbiology 2007 Vol.15 No.4 P.156-164
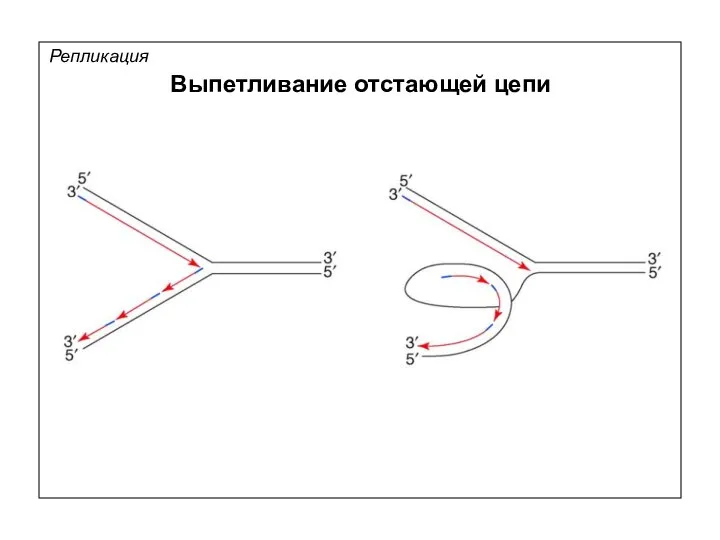
Репликация
Выпетливание отстающей цепи
Репликация
Выпетливание отстающей цепи
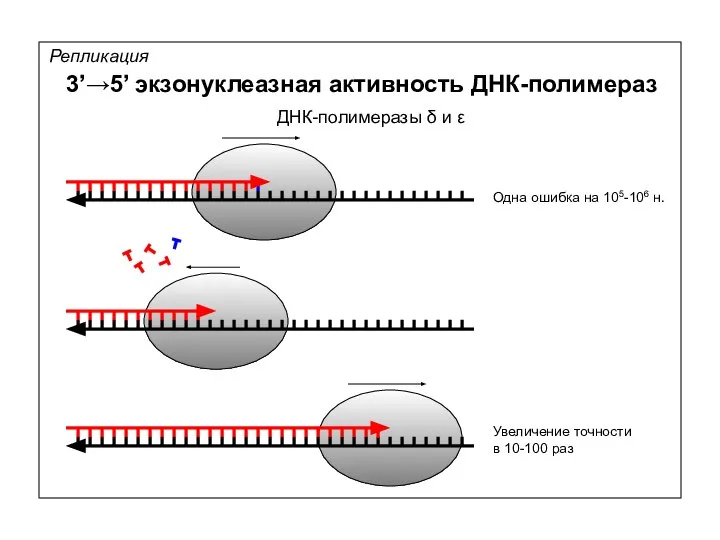
Репликация
3’→5’ экзонуклеазная активность ДНК-полимераз
Одна ошибка на 105-106 н.
Увеличение точности
в 10-100 раз
ДНК-полимеразы
Репликация
3’→5’ экзонуклеазная активность ДНК-полимераз
Одна ошибка на 105-106 н.
Увеличение точности
в 10-100 раз
ДНК-полимеразы

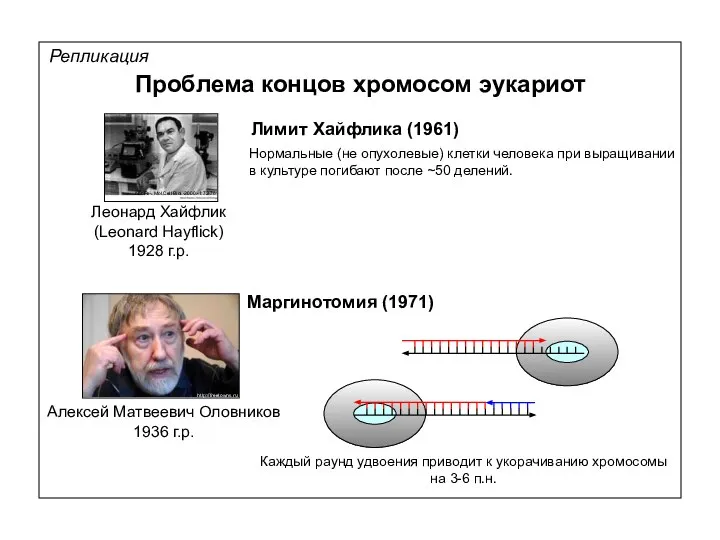
Репликация
Проблема концов хромосом эукариот
Маргинотомия (1971)
Лимит Хайфлика (1961)
Нормальные (не опухолевые) клетки человека
Репликация
Проблема концов хромосом эукариот
Маргинотомия (1971)
Лимит Хайфлика (1961)
Нормальные (не опухолевые) клетки человека
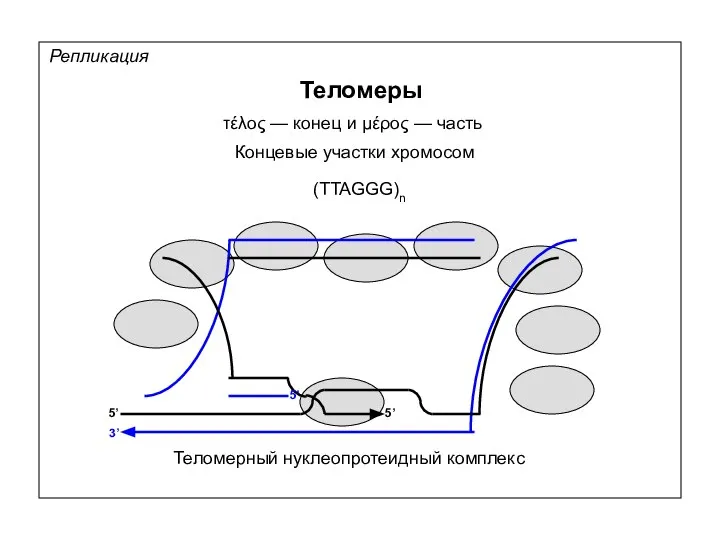
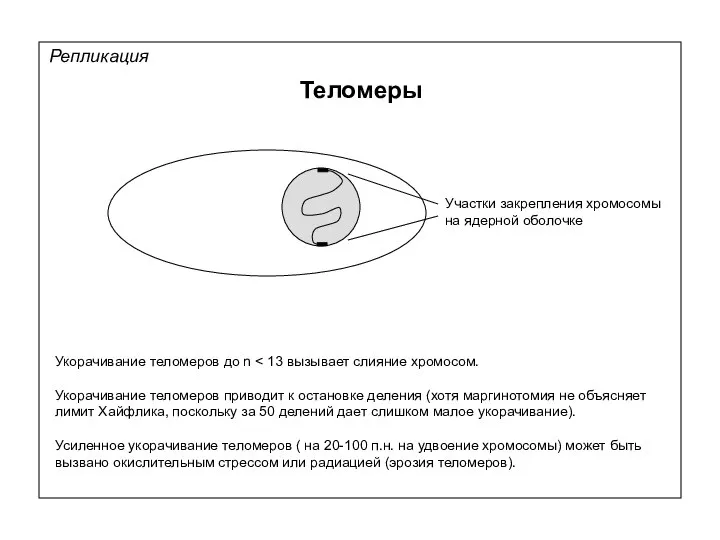
Репликация
Теломеры
τέλος — конец и μέρος — часть
Концевые участки хромосом
(TTAGGG)n
5’
5’
3’
5’
Теломерный нуклеопротеидный комплекс
Репликация
Теломеры
τέλος — конец и μέρος — часть
Концевые участки хромосом
(TTAGGG)n
5’
5’
3’
5’
Теломерный нуклеопротеидный комплекс
Репликация
Теломеры
Укорачивание теломеров до n < 13 вызывает слияние хромосом.
Укорачивание теломеров приводит
Репликация
Теломеры
Укорачивание теломеров до n < 13 вызывает слияние хромосом.
Укорачивание теломеров приводит
Репликация
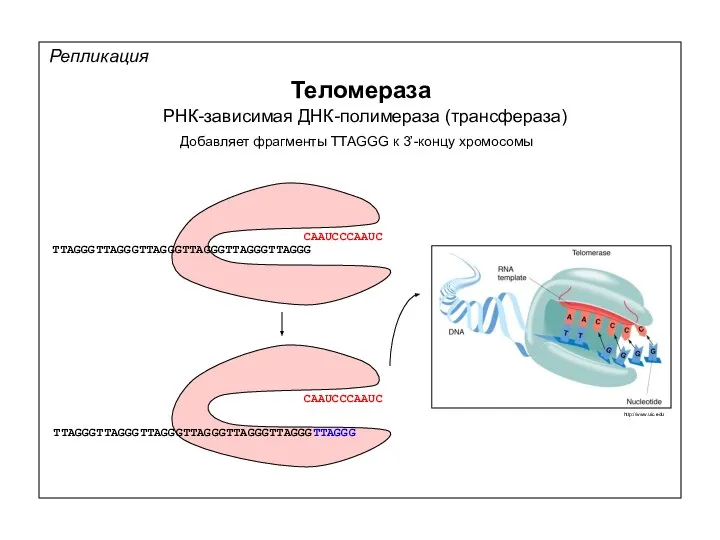
Теломераза
РНК-зависимая ДНК-полимераза (трансфераза)
Добавляет фрагменты TTAGGG к 3’-концу хромосомы
CAAUCCCAAUC
TTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGG
CAAUCCCAAUC
TTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGG
http://www.uic.edu
Репликация
Теломераза
РНК-зависимая ДНК-полимераза (трансфераза)
Добавляет фрагменты TTAGGG к 3’-концу хромосомы
CAAUCCCAAUC
TTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGG
CAAUCCCAAUC
TTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGG
http://www.uic.edu

Репликация
Теломераза
Нобелевская премия 2009 г.
за открытие механизмов защиты хромосом теломерами
и фермента теломеразы
Репликация
Теломераза
Нобелевская премия 2009 г.
за открытие механизмов защиты хромосом теломерами
и фермента теломеразы


Клеточный цикл
© Aaron Straight, Stanford University School of Medicine
Интерфаза (I)
Митоз
Клеточный цикл
© Aaron Straight, Stanford University School of Medicine
Интерфаза (I)
Митоз
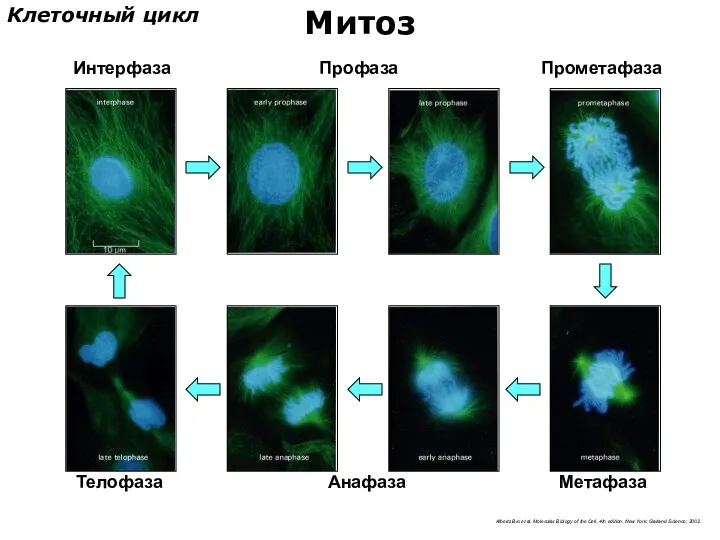
Интерфаза
Метафаза
Профаза
Прометафаза
Анафаза
Телофаза
Клеточный цикл
Митоз
Alberts Bю et al. Molecular Biology of the Cell. 4th
Интерфаза
Метафаза
Профаза
Прометафаза
Анафаза
Телофаза
Клеточный цикл
Митоз
Alberts Bю et al. Molecular Biology of the Cell. 4th
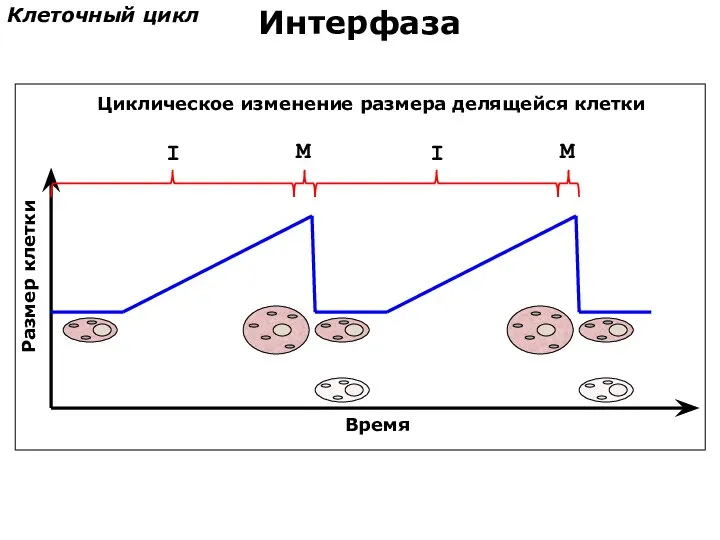
Размер клетки
Время
I
M
I
M
Клеточный цикл
Интерфаза
Циклическое изменение размера делящейся клетки
Размер клетки
Время
I
M
I
M
Клеточный цикл
Интерфаза
Циклическое изменение размера делящейся клетки
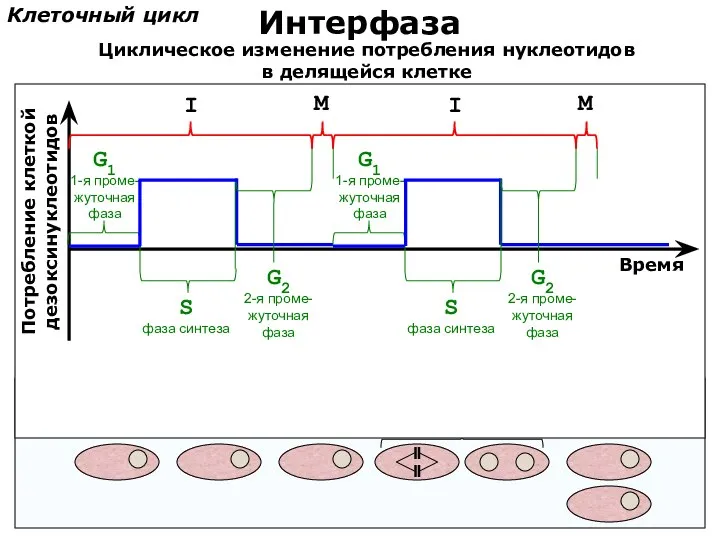
Клеточный цикл
Интерфаза
Потребление клеткой
дезоксинуклеотидов
Время
S
фаза синтеза
G1
1-я проме-
жуточная
фаза
G2
2-я проме-
жуточная
фаза
S
фаза синтеза
G1
1-я проме-
жуточная
фаза
G2
2-я проме-
жуточная
фаза
I
M
I
M
Циклическое изменение потребления
Клеточный цикл
Интерфаза
Потребление клеткой
дезоксинуклеотидов
Время
S
фаза синтеза
G1
1-я проме-
жуточная
фаза
G2
2-я проме-
жуточная
фаза
S
фаза синтеза
G1
1-я проме-
жуточная
фаза
G2
2-я проме-
жуточная
фаза
I
M
I
M
Циклическое изменение потребления
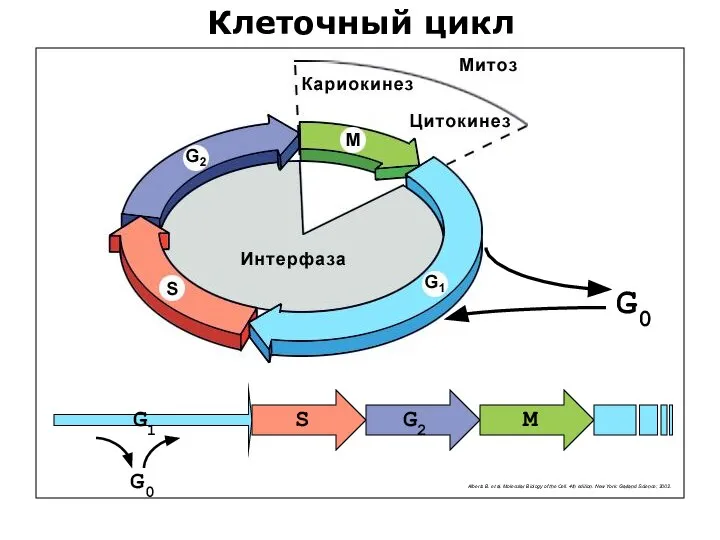
Клеточный цикл
G0
G1
G2
G0
S
M
Alberts B. et al. Molecular Biology of the Cell. 4th
Клеточный цикл
G0
G1
G2
G0
S
M
Alberts B. et al. Molecular Biology of the Cell. 4th
Клеточный цикл
Исследование клеточного цикла
http://www.cbp.pitt.edu/faculty/yong_wan/index.html
Клеточный цикл
Исследование клеточного цикла
http://www.cbp.pitt.edu/faculty/yong_wan/index.html
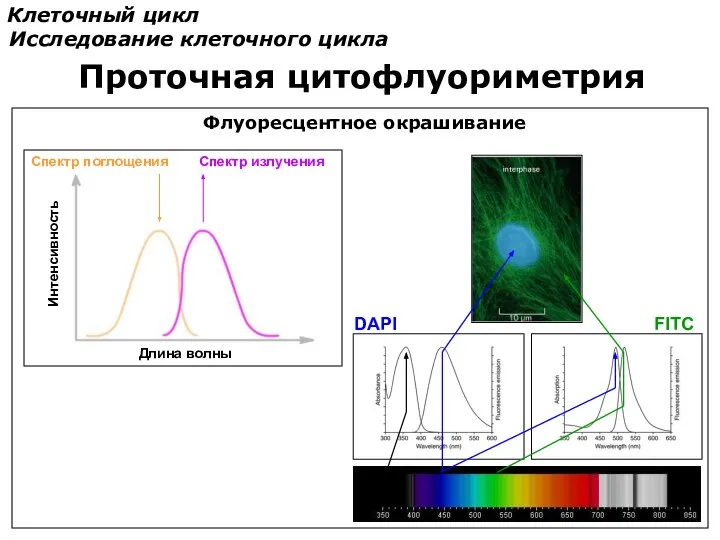
Клеточный цикл
Исследование клеточного цикла
Проточная цитофлуориметрия
Спектр поглощения
Спектр излучения
Длина волны
Интенсивность
DAPI
FITC
Флуоресцентное окрашивание
Клеточный цикл
Исследование клеточного цикла
Проточная цитофлуориметрия
Спектр поглощения
Спектр излучения
Длина волны
Интенсивность
DAPI
FITC
Флуоресцентное окрашивание
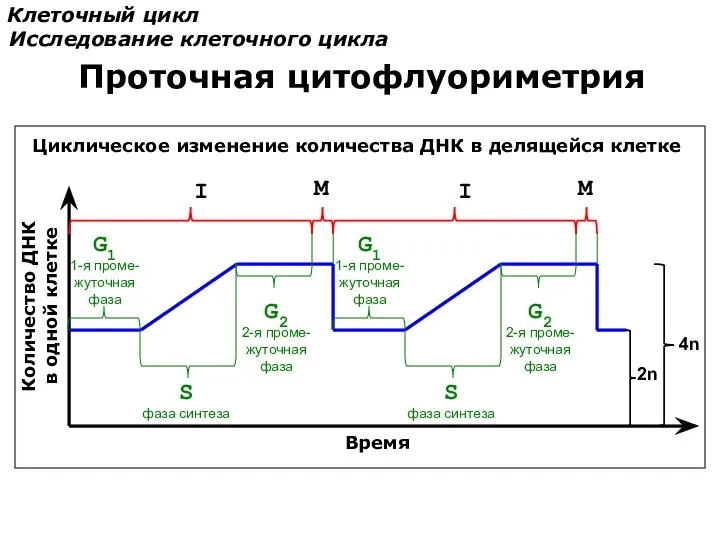
Количество ДНК
в одной клетке
Время
S
фаза синтеза
G1
1-я проме-
жуточная
фаза
G2
2-я проме-
жуточная
фаза
S
фаза синтеза
G1
1-я проме-
жуточная
фаза
G2
2-я проме-
жуточная
фаза
I
M
I
M
Клеточный цикл
Исследование
Количество ДНК
в одной клетке
Время
S
фаза синтеза
G1
1-я проме-
жуточная
фаза
G2
2-я проме-
жуточная
фаза
S
фаза синтеза
G1
1-я проме-
жуточная
фаза
G2
2-я проме-
жуточная
фаза
I
M
I
M
Клеточный цикл
Исследование
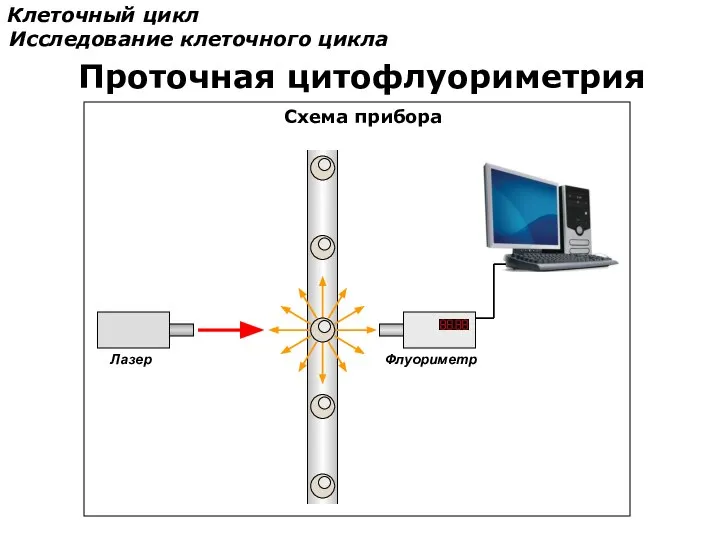
Клеточный цикл
Исследование клеточного цикла
Проточная цитофлуориметрия
Схема прибора
Клеточный цикл
Исследование клеточного цикла
Проточная цитофлуориметрия
Схема прибора
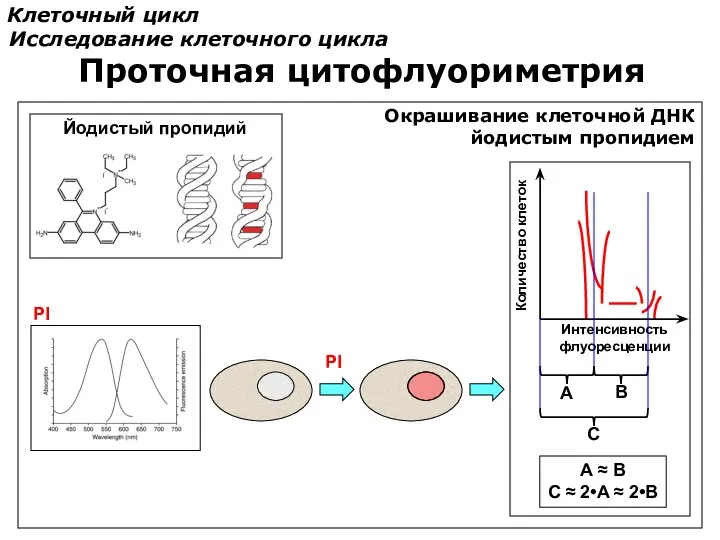
Клеточный цикл
Исследование клеточного цикла
Проточная цитофлуориметрия
Окрашивание клеточной ДНК
йодистым пропидием
PI
Интенсивность
флуоресценции
Количество клеток
A
B
C
A ≈ B
C
Клеточный цикл
Исследование клеточного цикла
Проточная цитофлуориметрия
Окрашивание клеточной ДНК
йодистым пропидием
PI
Интенсивность
флуоресценции
Количество клеток
A
B
C
A ≈ B
C
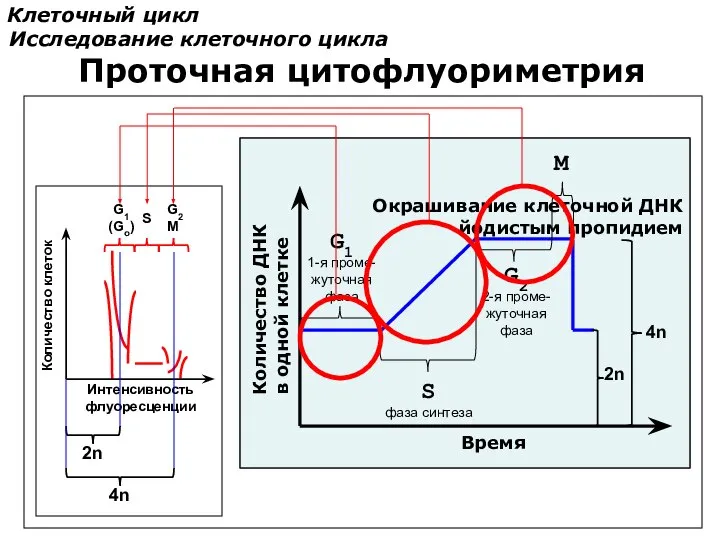
Клеточный цикл
Исследование клеточного цикла
Интенсивность
флуоресценции
Количество клеток
2n
4n
Окрашивание клеточной ДНК
йодистым пропидием
Количество ДНК
в одной клетке
Время
S
фаза
Клеточный цикл
Исследование клеточного цикла
Интенсивность
флуоресценции
Количество клеток
2n
4n
Окрашивание клеточной ДНК
йодистым пропидием
Количество ДНК
в одной клетке
Время
S
фаза
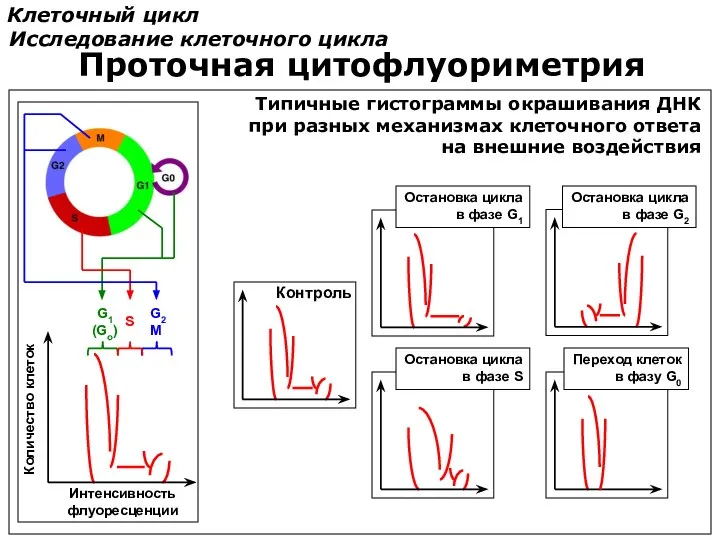
Клеточный цикл
Исследование клеточного цикла
Проточная цитофлуориметрия
Переход клеток
в фазу G0
Типичные гистограммы окрашивания ДНК
при
Клеточный цикл
Исследование клеточного цикла
Проточная цитофлуориметрия
Переход клеток
в фазу G0
Типичные гистограммы окрашивания ДНК
при


Репликация
Полимеразная цепная реакция (ПЦР)
Матрица
Симуляция работы хеликазы
Симуляция работы праймазы
Полимеризация ДНК
…
Репликация
Полимеразная цепная реакция (ПЦР)
Матрица
Симуляция работы хеликазы
Симуляция работы праймазы
Полимеризация ДНК
…
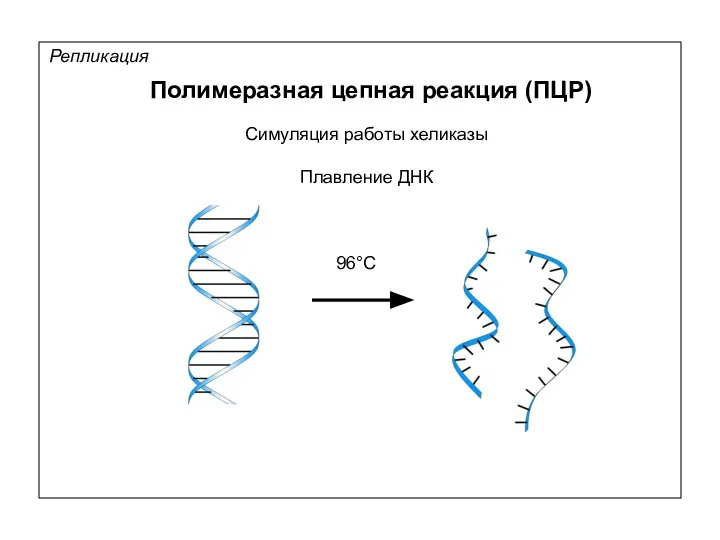
Репликация
Полимеразная цепная реакция (ПЦР)
Симуляция работы хеликазы
Плавление ДНК
96°С
Репликация
Полимеразная цепная реакция (ПЦР)
Симуляция работы хеликазы
Плавление ДНК
96°С

Репликация
Полимеразная цепная реакция (ПЦР)
Симуляция работы праймазы
Подбор и заказ праймеров
Отжиг праймеров
Репликация
Полимеразная цепная реакция (ПЦР)
Симуляция работы праймазы
Подбор и заказ праймеров
Отжиг праймеров

Репликация
Полимеразная цепная реакция (ПЦР)
Полимеризация ДНК
Thermus aquaticus
Wikimedia Commons
ДНК-полимераза Taq
Tопт = 72°С
Репликация
Полимеразная цепная реакция (ПЦР)
Полимеризация ДНК
Thermus aquaticus
Wikimedia Commons
ДНК-полимераза Taq
Tопт = 72°С

Репликация
ПЦР
1-й цикл
Плавление
Отжиг
Полимеризация
1 матрица (2 нити) + 1 полупродукт (2 нити)
Репликация
ПЦР
1-й цикл
Плавление
Отжиг
Полимеризация
1 матрица (2 нити) + 1 полупродукт (2 нити)

Репликация
ПЦР
2-й цикл
Плавление
Отжиг
Полимеризация
1 матрица (2 нити) + 2 полупродукта (4 нити) +
Репликация
ПЦР
2-й цикл
Плавление
Отжиг
Полимеризация
1 матрица (2 нити) + 2 полупродукта (4 нити) +

Репликация
ПЦР
3-й цикл
Плавление
Отжиг
Полимеризация
1 матрица (2 нити) + 3 полупродукта (6 нитей) +
Репликация
ПЦР
3-й цикл
Плавление
Отжиг
Полимеризация
1 матрица (2 нити) + 3 полупродукта (6 нитей) +

Репликация
ПЦР
Матриц - 1
Полупродуктов – n
Продуктов – 2n-(n+1)
Метод ПЦР позволяет определить наличие
Репликация
ПЦР
Матриц - 1
Полупродуктов – n
Продуктов – 2n-(n+1)
Метод ПЦР позволяет определить наличие
 Инсектициды на основе бакуловирусов
Инсектициды на основе бакуловирусов Полимер и пластмасса
Полимер и пластмасса Металлы в нашей жизни
Металлы в нашей жизни Сложные эфиры
Сложные эфиры Металлы и их соединения
Металлы и их соединения Властивості етанової (оцтової) кислоти
Властивості етанової (оцтової) кислоти Елементарний склад живих організмів
Елементарний склад живих організмів Структурно-механические свойства дисперсных систем
Структурно-механические свойства дисперсных систем Протеины: плюсы и минусы
Протеины: плюсы и минусы Излучение и поглощение света атомами. Виды спектров, спектральный анализ
Излучение и поглощение света атомами. Виды спектров, спектральный анализ Относительная атомная масса
Относительная атомная масса Биодизельное топливо
Биодизельное топливо Неметалічні елементи
Неметалічні елементи Бромды метил буының концентрациясын анықтау тәсілдемесі
Бромды метил буының концентрациясын анықтау тәсілдемесі Металлдардың кристалдық құрылымы
Металлдардың кристалдық құрылымы Поняття про лікарські препарати Підготували Стебко Марія, Бондар Аліна 11-Ф
Поняття про лікарські препарати Підготували Стебко Марія, Бондар Аліна 11-Ф  Закон постоянства состава веществ
Закон постоянства состава веществ Аргентум (Срібло)
Аргентум (Срібло) Мило. Синтетичні миючі засоби.
Мило. Синтетичні миючі засоби.  Растворы электролитов. Буферные растворы. Лекция 4
Растворы электролитов. Буферные растворы. Лекция 4 Аммиак. Физические и химические свойства. Получение и применение
Аммиак. Физические и химические свойства. Получение и применение Обмен железа и его нарушения
Обмен железа и его нарушения Презентация по Химии "Окислительно-восстановительные реакции" - скачать смотреть бесплатно
Презентация по Химии "Окислительно-восстановительные реакции" - скачать смотреть бесплатно Кетоны
Кетоны Альдегиды. Строение молекулы
Альдегиды. Строение молекулы Элементы третьего периода, периодической системы Д. И. Менделеева
Элементы третьего периода, периодической системы Д. И. Менделеева Нанохімія та нанотехнології
Нанохімія та нанотехнології Allgemeine Chemie für PharmazeutInnen
Allgemeine Chemie für PharmazeutInnen