- Кинетика микробиологических процессов

Содержание
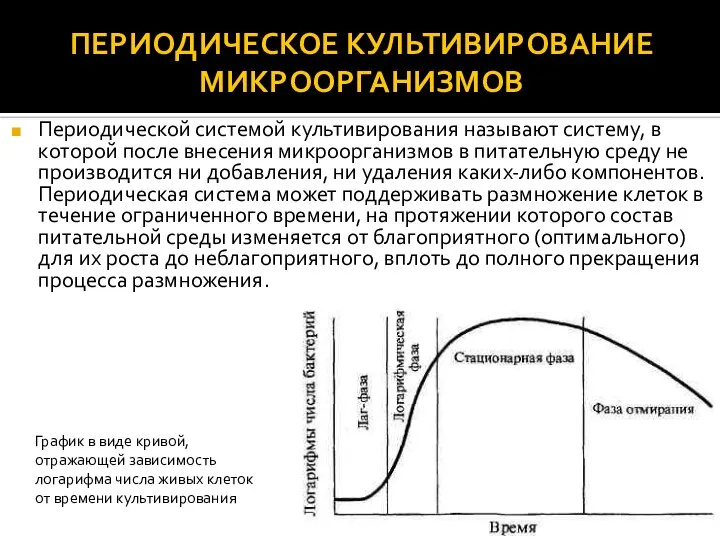
- 2. ПЕРИОДИЧЕСКОЕ КУЛЬТИВИРОВАНИЕ МИКРООРГАНИЗМОВ Периодической системой культивирования называют систему, в которой после внесения микроорганизмов в питательную среду
- 3. g = tудв = = µ = где dx – прирост биомассы за единицу времени =
- 4. Лимитирование микробного роста концентрацией субстрата заключается в следующем: чем выше концентрация субстрата, тем больше удельная скорость
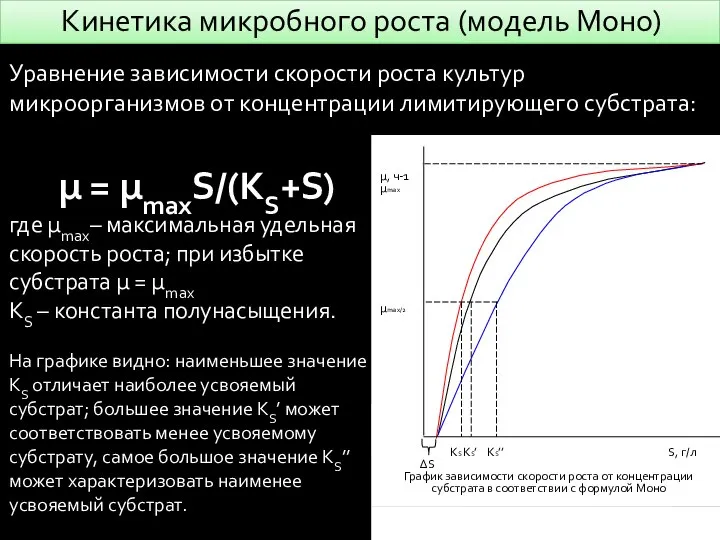
- 5. Кинетика микробного роста (модель Моно) µ = µmaxS/(KS+S) где µmax– максимальная удельная скорость роста; при избытке
- 6. Ингибирование и активация роста микроорганизмов Конкурентное ингибирование Обратимое ингибирование Неконкурентное ингибирование Необратимое ингибирование
- 7. Обратимое ингибирование Обратимые ингибиторы связываются с ферментом слабыми нековалентными связями и при определённых условиях легко отделяются
- 8. Обратимое ингибирование Неконкурентное ингибирование Неконкурентным называют такое ингибирование ферментативной реакции, при котором ингибитор взаимодействует с ферментом
- 9. Необратимое ингибирование Необратимое ингибирование наблюдают в случае образования ковалентных стабильных связей между молекулой ингибитора и фермента.
- 10. Ингибирование и активация роста микроорганизмов Полное конкурентное ингибирование В этом случае удельная скорость роста в экспоненциальной
- 11. Неконкурентное ингибирование Удельная скорость роста культуры в этом случае определяется формулой: μ = μmS[(Kr + βr)/(Kr
- 12. Влияние температуры на рост микроорганизмов Для каждого из микроорганизмов существует минимум, оптимум и максимум температурного режима
- 13. Уравнение зависимости роста от температуры При термообработке это число жизнеспособных клеток (N) микроорганизмов будет изменяться со
- 14. Низкие температуры замедляют рост микроорганизмов, но не убивают их. При температуре ниже 0 °С большинство микробов
- 15. Влияние рН на рост микроорганизмов 1) 2) Уравнение зависимости удельной скорости роста микроорганизмов от значений рН
- 16. Интегральная форма уравнения роста культуры микроорганизмов dX/dt = μX скорость роста μ(S) = μm · S/(Ks
- 17. Интегральная форма уравнения роста культуры микроорганизмов N = 1/A = YSo/Xo или Xm = Yso При
- 19. Скачать презентацию
ПЕРИОДИЧЕСКОЕ КУЛЬТИВИРОВАНИЕ МИКРООРГАНИЗМОВ
Периодической системой культивирования называют систему, в которой после внесения
ПЕРИОДИЧЕСКОЕ КУЛЬТИВИРОВАНИЕ МИКРООРГАНИЗМОВ
Периодической системой культивирования называют систему, в которой после внесения

g = tудв = =
µ =
где dx – прирост биомассы за
µ =
где dx – прирост биомассы за

Лимитирование микробного роста концентрацией субстрата заключается в следующем:
чем выше концентрация
Лимитирование микробного роста концентрацией субстрата заключается в следующем:
чем выше концентрация
Кинетика микробного роста (модель Моно)
µ = µmaxS/(KS+S)
где µmax– максимальная удельная
Кинетика микробного роста (модель Моно)
µ = µmaxS/(KS+S)
где µmax– максимальная удельная
Ингибирование и активация роста микроорганизмов
Конкурентное ингибирование
Обратимое ингибирование
Неконкурентное ингибирование
Необратимое ингибирование
Ингибирование и активация роста микроорганизмов
Конкурентное ингибирование
Обратимое ингибирование
Неконкурентное ингибирование
Необратимое ингибирование
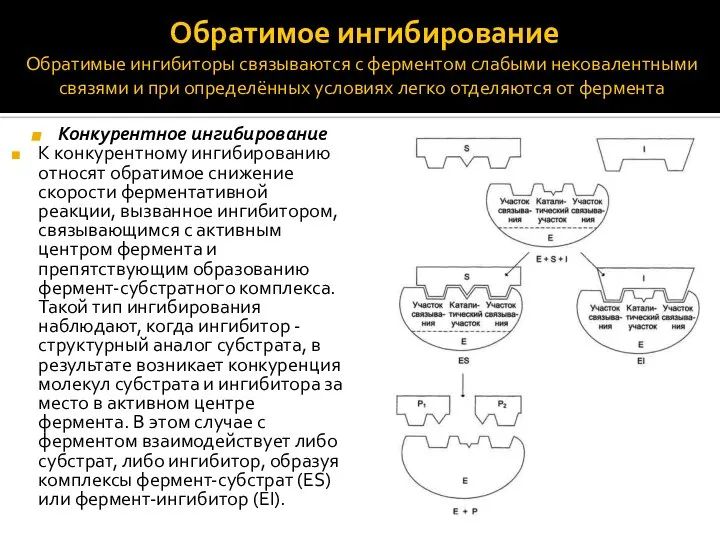
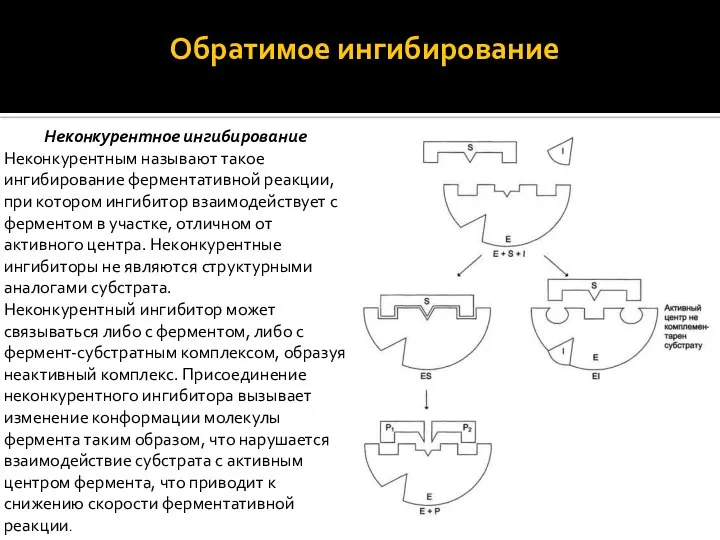
Обратимое ингибирование
Обратимые ингибиторы связываются с ферментом слабыми нековалентными связями и
Обратимое ингибирование Обратимые ингибиторы связываются с ферментом слабыми нековалентными связями и
Обратимое ингибирование
Неконкурентное ингибирование
Неконкурентным называют такое ингибирование ферментативной реакции, при котором
Обратимое ингибирование
Неконкурентное ингибирование
Неконкурентным называют такое ингибирование ферментативной реакции, при котором
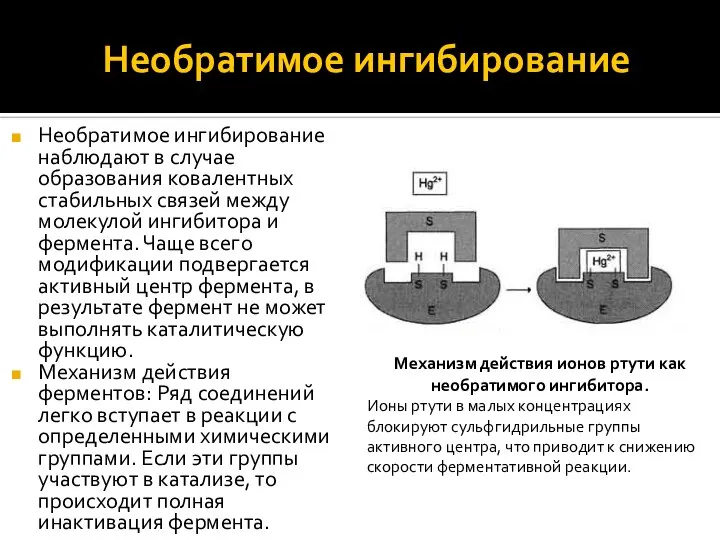
Необратимое ингибирование
Необратимое ингибирование наблюдают в случае образования ковалентных стабильных связей
Необратимое ингибирование
Необратимое ингибирование наблюдают в случае образования ковалентных стабильных связей
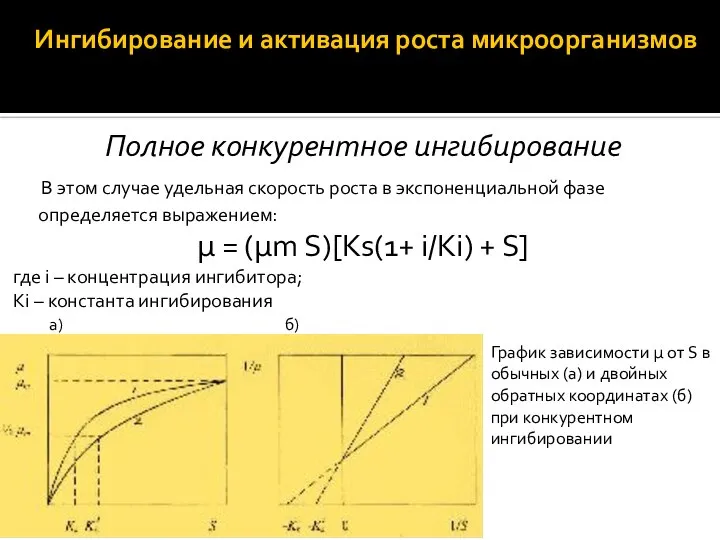
Ингибирование и активация роста микроорганизмов
Полное конкурентное ингибирование
В этом случае
Ингибирование и активация роста микроорганизмов
Полное конкурентное ингибирование
В этом случае
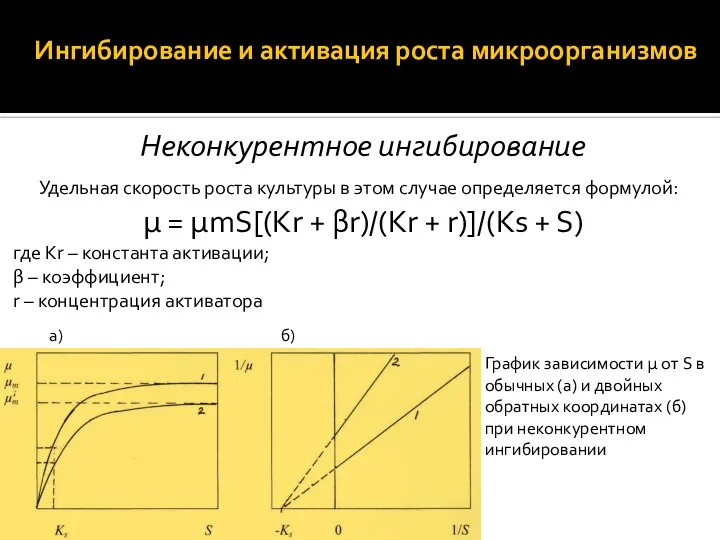
Неконкурентное ингибирование
Удельная скорость роста культуры в этом случае определяется формулой:
μ = μmS[(Kr + βr)/(Kr +
Неконкурентное ингибирование
Удельная скорость роста культуры в этом случае определяется формулой:
μ = μmS[(Kr + βr)/(Kr +
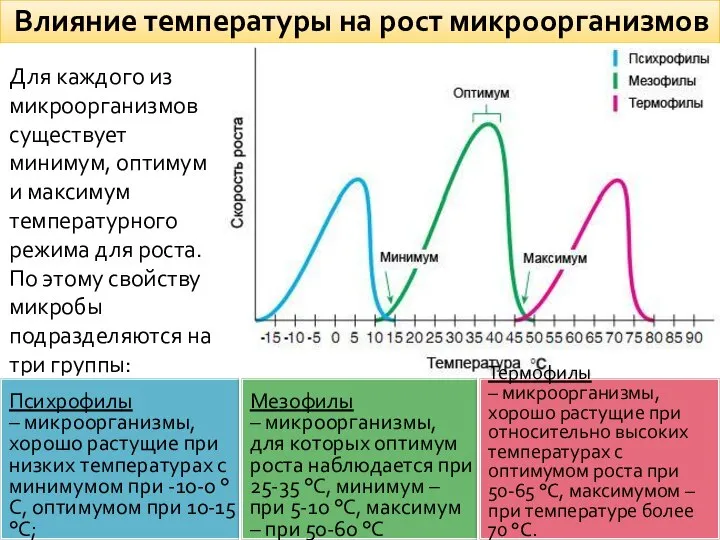
Влияние температуры на рост микроорганизмов
Для каждого из микроорганизмов существует минимум, оптимум
Влияние температуры на рост микроорганизмов
Для каждого из микроорганизмов существует минимум, оптимум
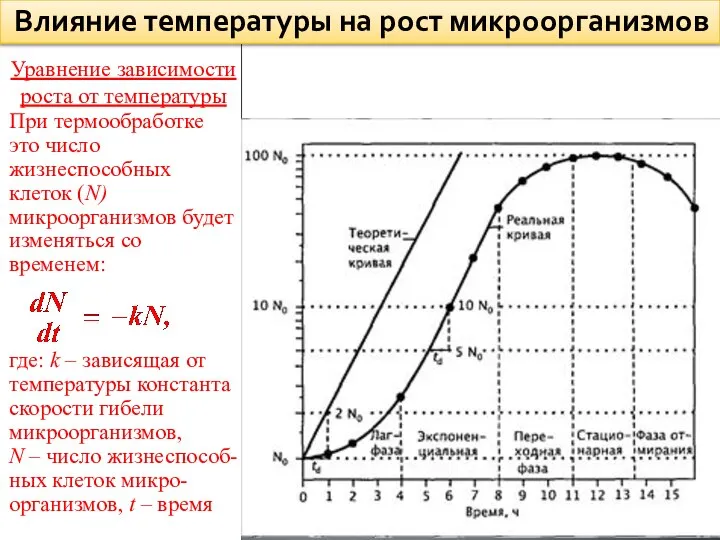
Уравнение зависимости роста от температуры
При термообработке это число жизнеспособных клеток (N)
Уравнение зависимости роста от температуры
При термообработке это число жизнеспособных клеток (N)

Низкие температуры замедляют рост микроорганизмов, но не убивают их. При температуре
Низкие температуры замедляют рост микроорганизмов, но не убивают их. При температуре
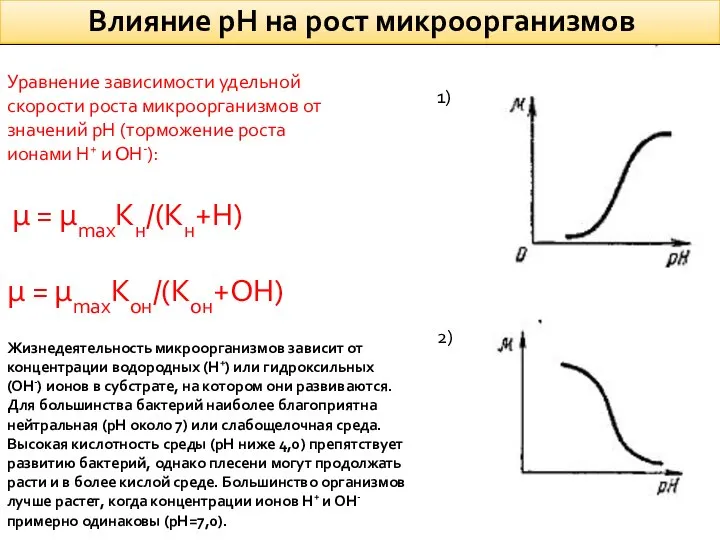
Влияние рН на рост микроорганизмов
1)
2)
Уравнение зависимости удельной скорости роста микроорганизмов от
Влияние рН на рост микроорганизмов
1)
2)
Уравнение зависимости удельной скорости роста микроорганизмов от

Интегральная форма уравнения роста культуры микроорганизмов
dX/dt = μX
скорость роста
μ(S) = μm · S/(Ks + S)
Уравнение Моно
Yср = (Х–
Интегральная форма уравнения роста культуры микроорганизмов
dX/dt = μX
скорость роста
μ(S) = μm · S/(Ks + S)
Уравнение Моно
Yср = (Х–

Интегральная форма уравнения роста культуры микроорганизмов
N = 1/A = YSo/Xo или Xm = Yso
При достаточно
Интегральная форма уравнения роста культуры микроорганизмов
N = 1/A = YSo/Xo или Xm = Yso
При достаточно
 Теория дискретных отображений
Теория дискретных отображений Что такое функция
Что такое функция Сравнение отрезков и углов
Сравнение отрезков и углов Фізико-математичний турнір «МІФ»
Фізико-математичний турнір «МІФ» Функция регрессии
Функция регрессии Аттестационная работа. Применение исследовательской и проектной деятельности в учебно-воспитательном процессе
Аттестационная работа. Применение исследовательской и проектной деятельности в учебно-воспитательном процессе Математика повсюду
Математика повсюду Путешествие в страну обыкновенных дробей
Путешествие в страну обыкновенных дробей История теоремы Пифагора, различные доказательства теоремы
История теоремы Пифагора, различные доказательства теоремы Вариационный ряд и его характеристии
Вариационный ряд и его характеристии Циклоидальные кривые
Циклоидальные кривые Окружность, хорды и дуги
Окружность, хорды и дуги Расчет установившихся режимов. Математические модели. Методы
Расчет установившихся режимов. Математические модели. Методы Кто хочет стать отличником. Павлова В.И. учитель математики СОГБОУ Духовщинской специальной /коррекционной/ общеобразователь
Кто хочет стать отличником. Павлова В.И. учитель математики СОГБОУ Духовщинской специальной /коррекционной/ общеобразователь Виды ДУ 1 порядка и методы их решения
Виды ДУ 1 порядка и методы их решения Многогранники
Многогранники Урок математики. 1 класс Тема: Числа от 1 до 20 Автор: Первушина Наталья Михайловна учитель нача
Урок математики. 1 класс Тема: Числа от 1 до 20 Автор: Первушина Наталья Михайловна учитель нача Понятие логарифма
Понятие логарифма Прямоугольный треугольник
Прямоугольный треугольник Оператор примитивной рекурсии
Оператор примитивной рекурсии Из истории параллельности прямых на плоскости
Из истории параллельности прямых на плоскости Екі факторлы дисперсиялық талдау
Екі факторлы дисперсиялық талдау Линейные уравнения с одной переменной
Линейные уравнения с одной переменной Число 2. Слева, справа, на, под. Толстый, тонкий
Число 2. Слева, справа, на, под. Толстый, тонкий Интерактивный тренажер «Показательные и логарифмические уравнения»
Интерактивный тренажер «Показательные и логарифмические уравнения» Погрешности приближённых вычислений. (Лекции 1-2)
Погрешности приближённых вычислений. (Лекции 1-2) Презентация по математике "Тиждень математики" - скачать
Презентация по математике "Тиждень математики" - скачать  Свойства действий с рациональными числами. Часть 2
Свойства действий с рациональными числами. Часть 2