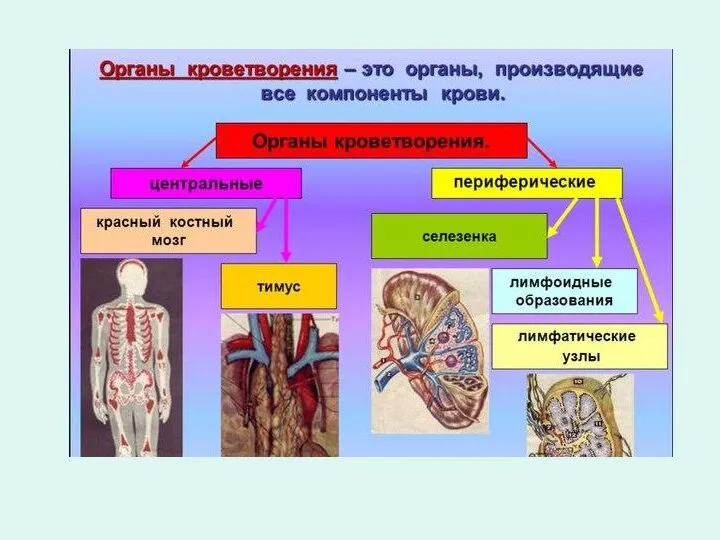
- Гемопоэз. Органы кроветворения

Содержание
- 2. гемопоэз Более 30 млн клеток в минуту образуется в кроветворных органах, а в течение жизни человека
- 3. гемопоэз Кроветворение (гемопоэз) - многостадийный процесс дифференцировки клеточных элементов, в результате которого образуются эритроциты, лейкоциты, тромбоциты,
- 5. Клетки, способные восстанавливать гемопоэз после облучения или токсических воздействий, носят название «стволовых клеток». Впервые термин «стволовые
- 6. «Из стволовых клеток можно вырастить любой орган ?» Орган вырастить нельзя но клетки и ткани –вполне
- 7. способность к самообновлению способность к дифференцировке количество стволовых клеток ограничено Источники: Костный мозг Пуповинная кровь Периферическая
- 8. Для лечения лейкозов и других заболеваний крови При лечении рассеянного склероза Стволовые клетки кожи используются при
- 9. Стволовые клетки Кроветорные СК Эмбриональные СК Стромальные (индуцированные) СК
- 10. Стволовые клетки взрослого организма мультипотентны или унипотентны. А эмбриональные стволовые клетки плюрипотентны. Стволовые клетки по настоящему
- 11. Уникальное свойство эмбриональных стволовых клеток — тотипотентность Проблема №1: как направить их по пути дифференцировки Проблема
- 12. Костный мозг состоит из двух видов стволовых клеток: гемопоэтические стволовые клетки стромальные стволовые клетки У новорожденного
- 13. Стромальные стволовые клетки костного мозга - универсальны. Они поступают с кровотоком в поврежденный орган или ткань
- 14. Выделены уникальные вещества: особые белки, вызывающие перерождение стромальных клеток в клетки костной ткани (остеобласты). В США
- 15. В 2012 г. в Стокгольме объявили лауреатов Нобелевской премии по физиологии и медицине. Награда присуждена Джону
- 16. Гемопоэз поддерживается в течение всей жизни за счёт «стволовых клеток» Нормальное кроветворение поликлональное, т. е. осуществляется
- 18. Регуляция гемопоэза Позитивные: КСФ, ИЛ-6, ИЛ-11, ИЛ-12 Негативные: TGF-β, MIP-1α, ФНО-α, интерферон-α, интерферон-γ, кислые изоферритины, лактоферрин
- 19. Регуляция гемопоэза 1. Факторы, влияющие на ранние СКК: фактор стволовых клеток (ФСК), гранулоцитарный колониестимулирующий фактор (Г-КСФ),
- 20. Последовательность ультраструктурных изменений в процессе некроза и апоптоза
- 21. Молекулярные механизмы апоптоза. Выделяют два типа сигнальных путей, вызывающих апоптоз, - повреждение ДНК, радиация и другие
- 22. Бурстстимулиру-ющая активность Витамин В12 , фолиевая кислота, железо, медь Эритропоэз
- 23. Иммунологический маркёр эритробластов – гликофорин А Дифференцировка и созревание эритроидных клеток от проэритробласта до эритроцита -
- 24. Синтез гемоглобина начинается на стадии проэритробласта В норме ранний полихроматофильный эритробласт подходит к последнему митозу с
- 25. Проэритробласт (эритробласт) Пронормобласт
- 26. Оксифильный эритробласт Базофильный (1) и полихрома- тофильный (2) эритробласты
- 27. Ретикулоциты Эритроциты
- 28. Распределение популяции ретикулоцитов по степени зрелости
- 29. Диаметр – 7,2-8,0 мкм Площадь поверхности – 140мкм2 Объём – 90мкм3 Нормохромные Нормоцит-нормальный эритроцит
- 30. Эритроциты в капилляре существенно меняют форму за счет деформируемости мембраны, что позволяет им проходить сквозь узкие
- 31. Схема мембраны эритроцита
- 32. Функции эритроцитов Участие в газообмене и поддержании буферного кислотно-основного равновесия крови Определяют реологию крови Участвуют в
- 33. Гранулоцитопоэз В процессе созревания (в течение 10-13 дней) происходит: Уменьшение ядра Конденсация хроматина Исчезновение ядрышек Сегментация
- 34. Гранулогенез Первичные гранулы – азурофильные. Маркёр – миелопероксидаза. Содержат: лизоцим, катепсин, эластазу, миелопероксидазу, антибактериальные катионные белки,
- 35. Миелоидный росток Миелобласт Промиелоцит
- 36. Клетки нейтрофильного ростка
- 37. сегментоядерные нейтрофилы, Тельце Барра (1)
- 38. Клеточно-гуморальные контакты нейтрофилов
- 39. Основные этапы фагоцитоза – хемотаксис, адгезия антигена к рецепторам мембраны фагоцита, погружение в цитоплазму (эндоцитоз), образование
- 40. Эозинофилы На мембране есть рецепторы к Fc-фрагменту иммуноглобулинов, рецепторы для компонентов комплемента, молекулы адгезии. CD52,CD69, CD40
- 41. Палочкоядерный эозинофил эозинофил
- 42. Базофил Тучная клетка
- 43. Базофилы В гранулах: гистамин, гистидин, гепарин, серотонин, ферменты, лейкотриены, тромбоксаны, простогландины, фактор хемотаксиса эозинофилов, фактор активации
- 44. Участие тучных клеток в реакции гиперчувствительности немедленного типа. ПГ - простагландины, ЛТ - лейкотриены, ЛТВ4 -
- 45. Моноцитопоэз. Моноцитарный росток Монобласт Промоноцит
- 46. Моноцит Макрофаг
- 47. Цитохимические маркёры клеток моноцитарно-макрофагальной линии Неспецифическая эстераза, подавляемая фторидом натрия Кислая фосфатаза В процессе дифференцировки уменьшается
- 48. Мононуклеарные фагоциты Обновление тканевых макрофагов происходит за счёт притока моноцитов крови 56% - макрофаги печени (клетки
- 49. Презентация антигена антигенпредставляющими клетками Макрофаги разделены на 2 касса: Антигенперерабатывающие (профессиональные фагоциты) Антигенпредставляющие (дендритные клетки)
- 50. Фолликулярные дендритные клетки Локализация: В В-клеточных зонах лимфоузлов В селезёнке В мукозо-ассоциированной лимфоидной ткани (MALT) Представляют
- 51. Интердигитальные дендритные клетки Локализация: В парокортикальных зонах лимфоузлов В Т-зависимых зонах селезёнки, миндалин В мозговом веществе
- 52. Биологическая роль СМФ в процессах обмена веществ Участие в углеводном обмене (рецепторы к инсулину и способность
- 53. Секреторные факторы макрофагов Провоспалительные цитокины (ИЛ-1, ИЛ-6, ИЛ-8, ИЛ-12, ИЛ-18, ФНО-α, интерферон-α, моноцитарный хемотаксический протеин, миграцию
- 54. Роль цитокинов в воспалении. Активация эндотелиальных клеток, повышение экспрессии молекул адгезии - Е-селектин, Р-селектин, активация нейтрофилов,
- 55. Лимфоцитопоэз. Лимфоцитарный росток лимфобласт пролимфоцит лимфоцит
- 56. Большой гранулярный лимфоцит Плазмобласт
- 57. Проплазмоцит Плазмотическая клетка
- 58. Антигеннезависимые и антигензависимые этапы дифференцировки Т-лим- фоцитов
- 60. Дифференцировка В-лимфоцитов Основной характеристикой В-лимфоцитов является наличие на их мембране рецепторов для распознавания антигенов, основу которых
- 61. Механизм гуморального иммунитета: Интерферон Лизоцим +
- 63. Мегакариоцитарный росток мегакариобласт промегакариоцит мегакариоцит
- 64. Активированные тромбоциты на поврежденной сосудистой стенке. При активации происходит округление тромбоцитов и образование псевдоподий, с помощью
- 66. Скачать презентацию

гемопоэз
Более 30 млн клеток в минуту образуется в кроветворных органах, а
гемопоэз
Более 30 млн клеток в минуту образуется в кроветворных органах, а

гемопоэз
Кроветворение (гемопоэз) - многостадийный процесс дифференцировки клеточных элементов, в результате которого
гемопоэз
Кроветворение (гемопоэз) - многостадийный процесс дифференцировки клеточных элементов, в результате которого

Клетки, способные восстанавливать гемопоэз после облучения или токсических воздействий, носят название
Клетки, способные восстанавливать гемопоэз после облучения или токсических воздействий, носят название
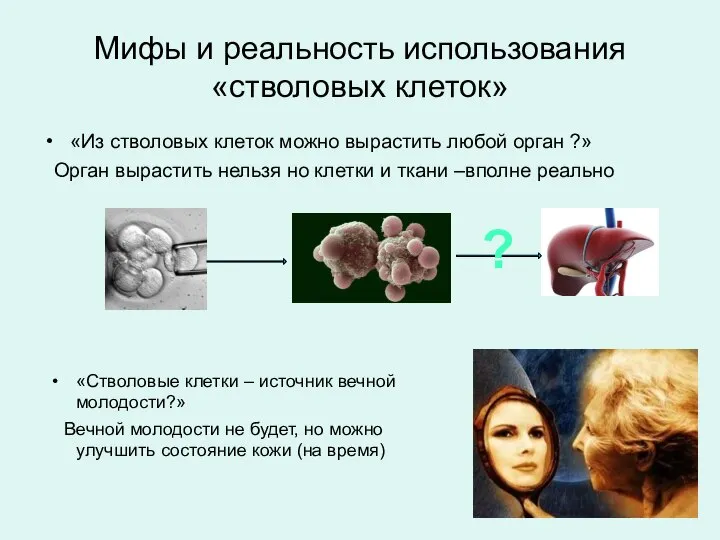
«Из стволовых клеток можно вырастить любой орган ?»
Орган вырастить нельзя
«Из стволовых клеток можно вырастить любой орган ?»
Орган вырастить нельзя

способность к самообновлению
способность к дифференцировке
количество стволовых клеток ограничено
Источники:
Костный мозг
Пуповинная
способность к самообновлению
способность к дифференцировке
количество стволовых клеток ограничено
Источники:
Костный мозг
Пуповинная

Для лечения лейкозов и других заболеваний крови
При лечении рассеянного склероза
Стволовые
Для лечения лейкозов и других заболеваний крови
При лечении рассеянного склероза
Стволовые

Стволовые клетки
Кроветорные СК
Эмбриональные СК
Стромальные (индуцированные) СК
Стволовые клетки
Кроветорные СК
Эмбриональные СК
Стромальные (индуцированные) СК
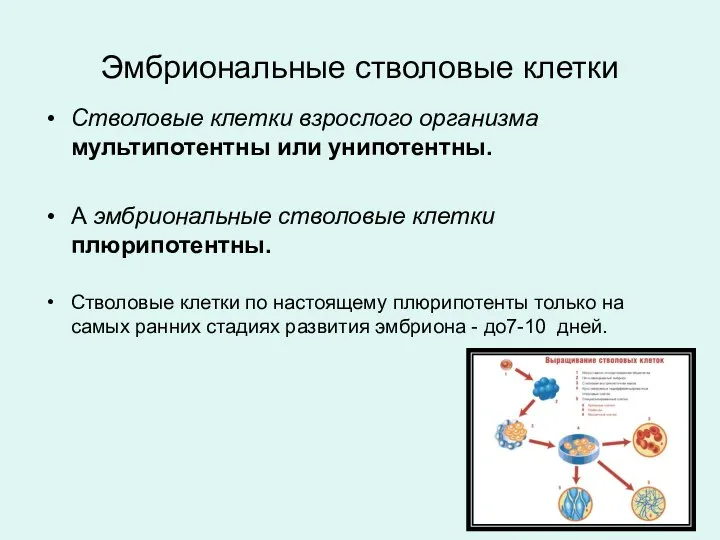
Стволовые клетки взрослого организма мультипотентны или унипотентны.
А эмбриональные стволовые клетки плюрипотентны.
Стволовые
Стволовые клетки взрослого организма мультипотентны или унипотентны.
А эмбриональные стволовые клетки плюрипотентны.
Стволовые

Уникальное свойство эмбриональных стволовых клеток — тотипотентность
Проблема №1: как направить их
Уникальное свойство эмбриональных стволовых клеток — тотипотентность
Проблема №1: как направить их

Костный мозг состоит из двух видов стволовых клеток:
гемопоэтические стволовые клетки
стромальные
Костный мозг состоит из двух видов стволовых клеток:
гемопоэтические стволовые клетки
стромальные
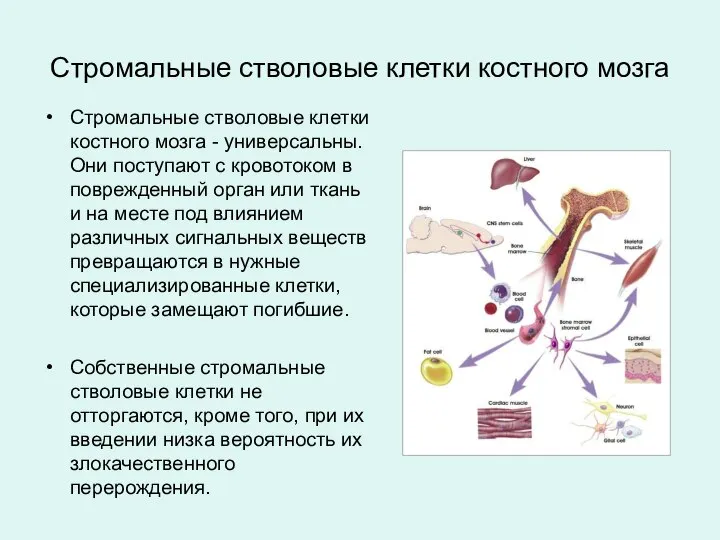
Стромальные стволовые клетки костного мозга - универсальны. Они поступают с кровотоком
Стромальные стволовые клетки костного мозга - универсальны. Они поступают с кровотоком

Выделены уникальные вещества: особые белки, вызывающие перерождение стромальных клеток в клетки
Выделены уникальные вещества: особые белки, вызывающие перерождение стромальных клеток в клетки

В 2012 г. в Стокгольме объявили лауреатов Нобелевской премии по физиологии
В 2012 г. в Стокгольме объявили лауреатов Нобелевской премии по физиологии

Гемопоэз поддерживается в течение всей жизни за счёт «стволовых клеток»
Нормальное кроветворение
Гемопоэз поддерживается в течение всей жизни за счёт «стволовых клеток»
Нормальное кроветворение

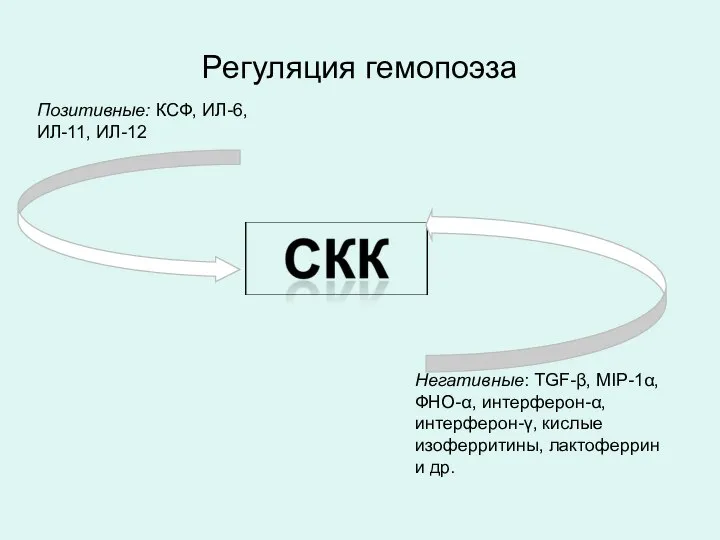
Регуляция гемопоэза
Позитивные: КСФ, ИЛ-6, ИЛ-11, ИЛ-12
Негативные: TGF-β, MIP-1α, ФНО-α, интерферон-α, интерферон-γ,
Регуляция гемопоэза
Позитивные: КСФ, ИЛ-6, ИЛ-11, ИЛ-12
Негативные: TGF-β, MIP-1α, ФНО-α, интерферон-α, интерферон-γ,

Регуляция гемопоэза
1. Факторы, влияющие на ранние СКК: фактор стволовых клеток (ФСК),
Регуляция гемопоэза
1. Факторы, влияющие на ранние СКК: фактор стволовых клеток (ФСК),
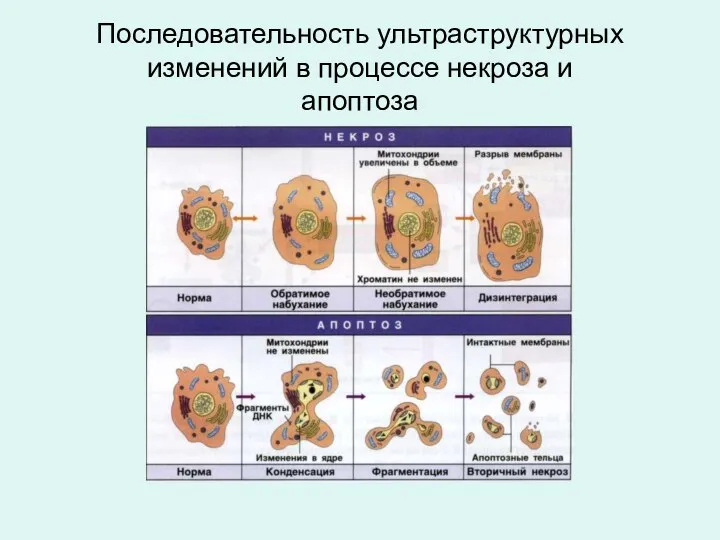
Последовательность ультраструктурных изменений в процессе некроза и
апоптоза
Последовательность ультраструктурных изменений в процессе некроза и
апоптоза

Молекулярные механизмы апоптоза.
Выделяют два типа сигнальных путей, вызывающих апоптоз, -
Молекулярные механизмы апоптоза.
Выделяют два типа сигнальных путей, вызывающих апоптоз, -
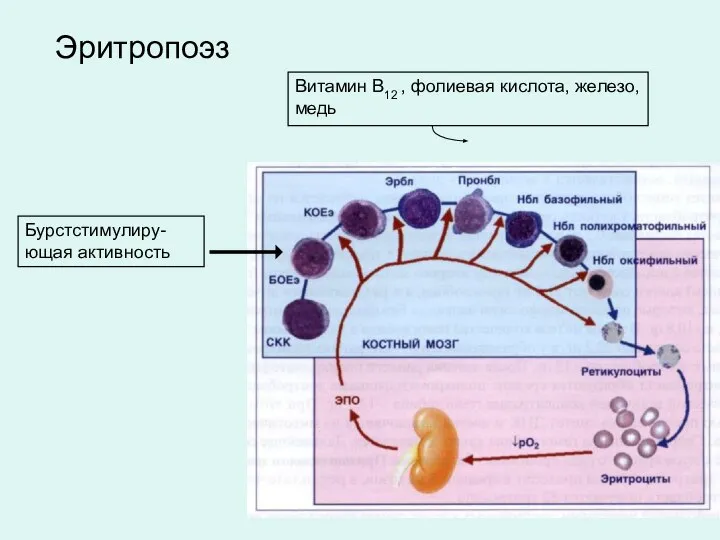
Бурстстимулиру-ющая активность
Витамин В12 , фолиевая кислота, железо, медь
Эритропоэз
Бурстстимулиру-ющая активность
Витамин В12 , фолиевая кислота, железо, медь
Эритропоэз

Иммунологический маркёр эритробластов – гликофорин А
Дифференцировка и созревание эритроидных клеток от
Иммунологический маркёр эритробластов – гликофорин А
Дифференцировка и созревание эритроидных клеток от

Синтез гемоглобина начинается на стадии проэритробласта
В норме ранний полихроматофильный эритробласт подходит
Синтез гемоглобина начинается на стадии проэритробласта
В норме ранний полихроматофильный эритробласт подходит
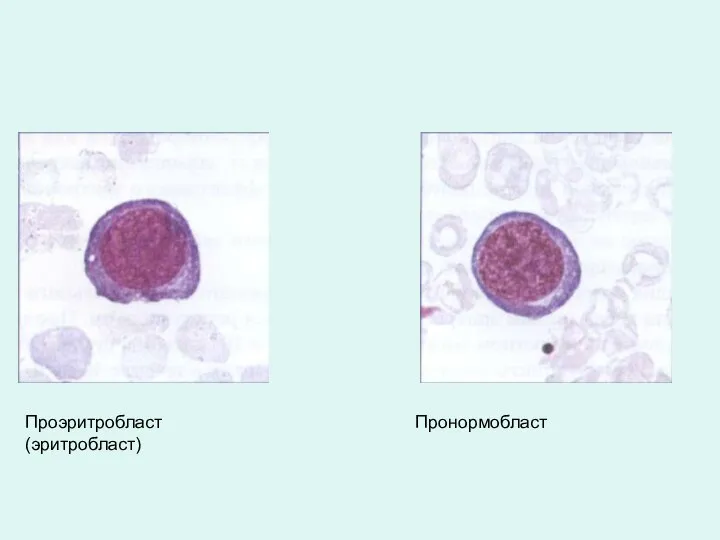
Проэритробласт (эритробласт)
Пронормобласт
Проэритробласт (эритробласт)
Пронормобласт

Оксифильный эритробласт
Базофильный (1) и полихрома- тофильный (2) эритробласты
Оксифильный эритробласт
Базофильный (1) и полихрома- тофильный (2) эритробласты
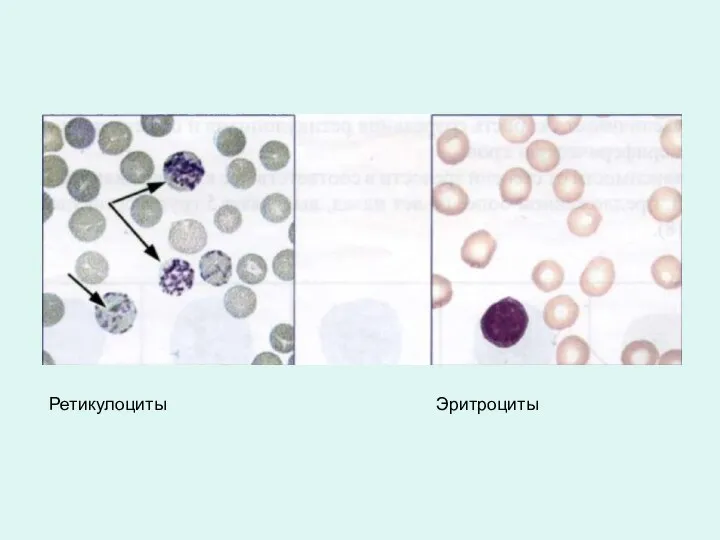
Ретикулоциты Эритроциты
Ретикулоциты Эритроциты
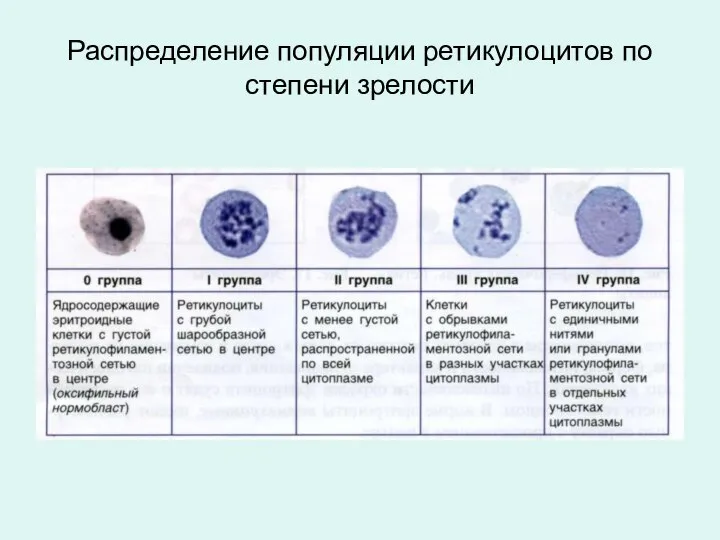
Распределение популяции ретикулоцитов по степени зрелости
Распределение популяции ретикулоцитов по степени зрелости
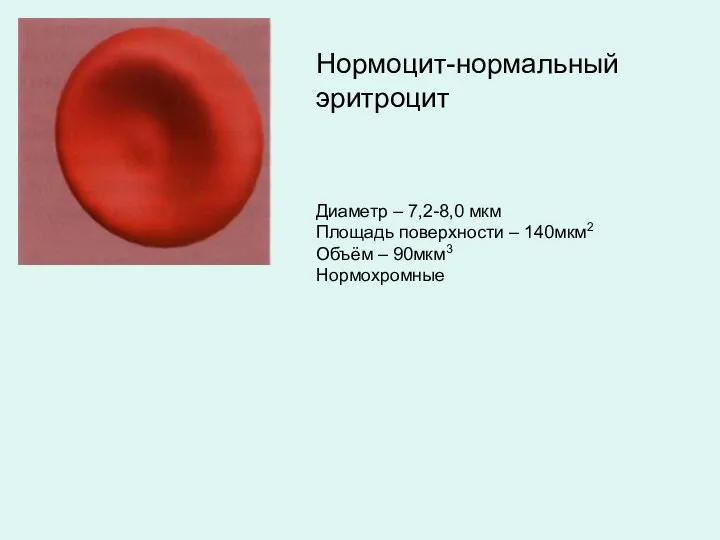
Диаметр – 7,2-8,0 мкм
Площадь поверхности – 140мкм2
Объём – 90мкм3
Нормохромные
Нормоцит-нормальный эритроцит
Диаметр – 7,2-8,0 мкм
Площадь поверхности – 140мкм2
Объём – 90мкм3
Нормохромные
Нормоцит-нормальный эритроцит

Эритроциты в капилляре существенно
меняют форму за счет деформируемости
мембраны, что позволяет им
проходить
Эритроциты в капилляре существенно
меняют форму за счет деформируемости
мембраны, что позволяет им
проходить
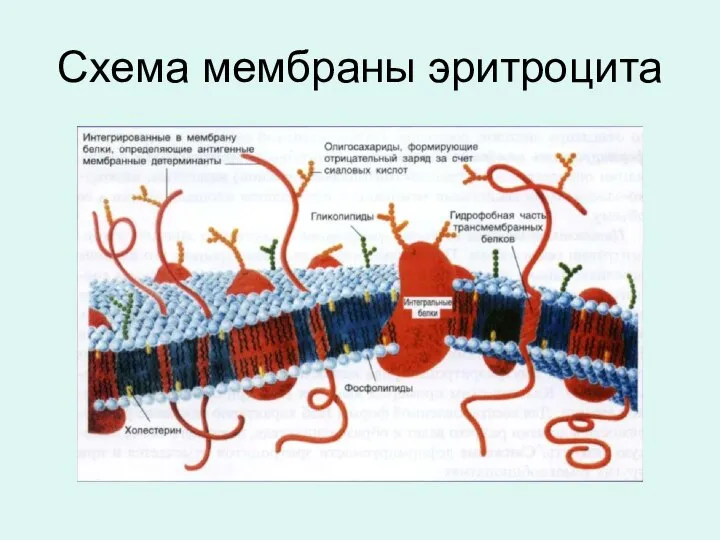
Схема мембраны эритроцита
Схема мембраны эритроцита

Функции эритроцитов
Участие в газообмене и поддержании буферного кислотно-основного равновесия крови
Определяют реологию
Функции эритроцитов
Участие в газообмене и поддержании буферного кислотно-основного равновесия крови
Определяют реологию

Гранулоцитопоэз
В процессе созревания (в течение 10-13 дней) происходит:
Уменьшение ядра
Конденсация хроматина
Исчезновение
Гранулоцитопоэз
В процессе созревания (в течение 10-13 дней) происходит:
Уменьшение ядра
Конденсация хроматина
Исчезновение

Гранулогенез
Первичные гранулы – азурофильные. Маркёр – миелопероксидаза. Содержат: лизоцим, катепсин,
Гранулогенез
Первичные гранулы – азурофильные. Маркёр – миелопероксидаза. Содержат: лизоцим, катепсин,
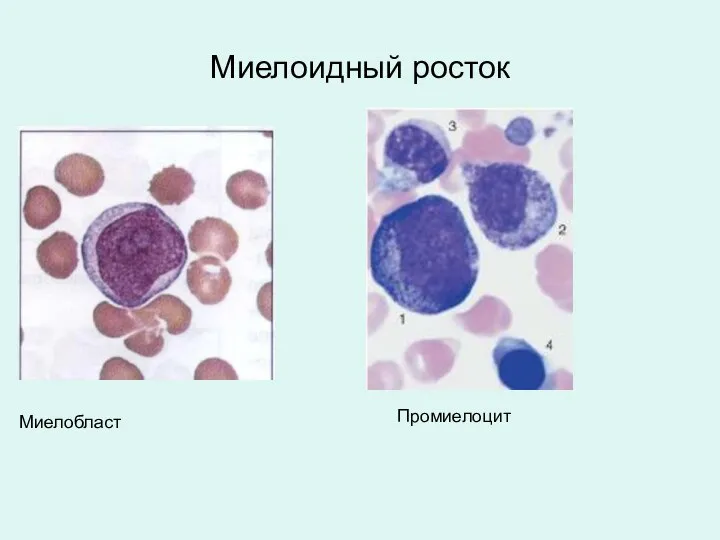
Миелоидный росток
Миелобласт
Промиелоцит
Миелоидный росток
Миелобласт
Промиелоцит

Клетки нейтрофильного ростка
Клетки нейтрофильного ростка

сегментоядерные нейтрофилы,
Тельце Барра (1)
сегментоядерные нейтрофилы,
Тельце Барра (1)
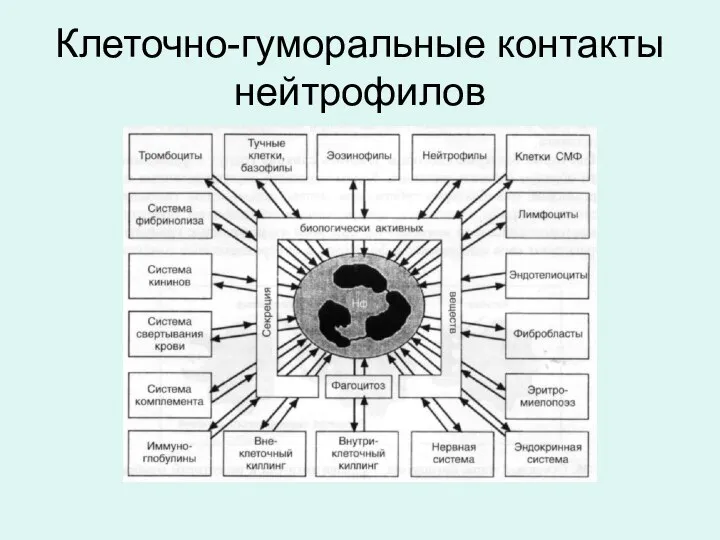
Клеточно-гуморальные контакты нейтрофилов
Клеточно-гуморальные контакты нейтрофилов
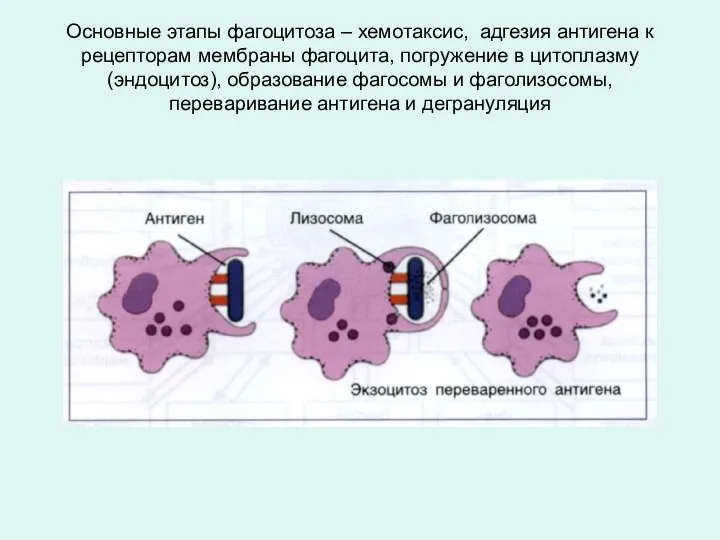
Основные этапы фагоцитоза – хемотаксис, адгезия антигена к рецепторам мембраны фагоцита,
Основные этапы фагоцитоза – хемотаксис, адгезия антигена к рецепторам мембраны фагоцита,

Эозинофилы
На мембране есть рецепторы к Fc-фрагменту иммуноглобулинов, рецепторы для компонентов
Эозинофилы
На мембране есть рецепторы к Fc-фрагменту иммуноглобулинов, рецепторы для компонентов
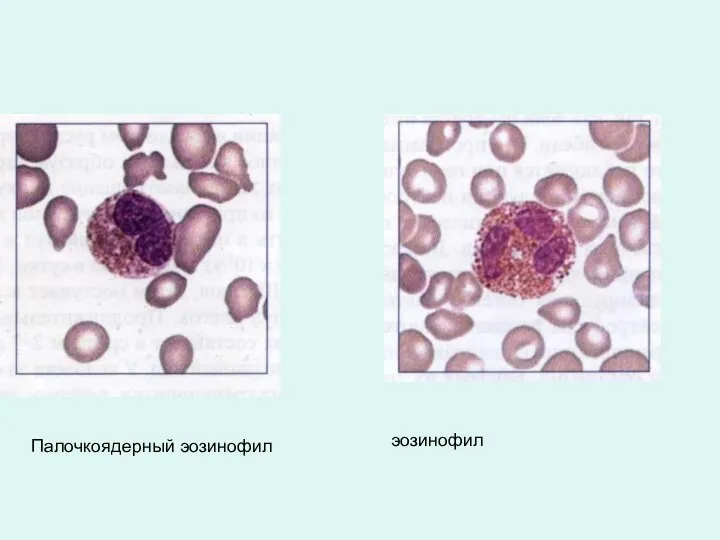
Палочкоядерный эозинофил
эозинофил
Палочкоядерный эозинофил
эозинофил
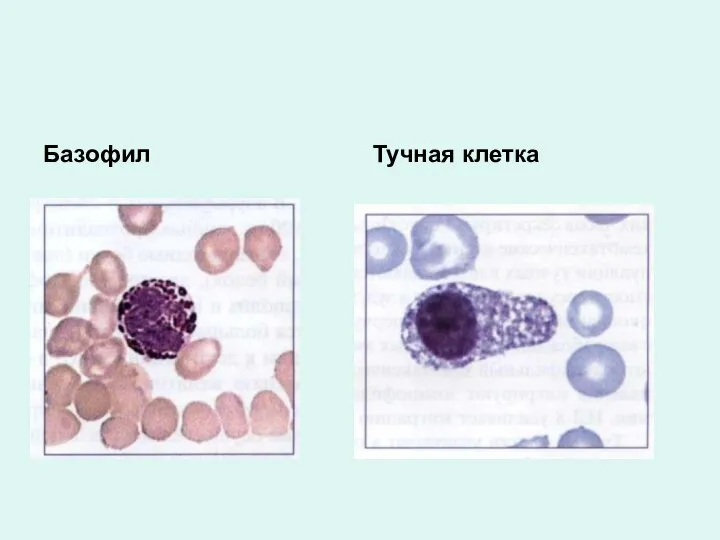
Базофил
Тучная клетка
Базофил
Тучная клетка

Базофилы
В гранулах: гистамин, гистидин, гепарин, серотонин, ферменты, лейкотриены, тромбоксаны, простогландины,
Базофилы
В гранулах: гистамин, гистидин, гепарин, серотонин, ферменты, лейкотриены, тромбоксаны, простогландины,
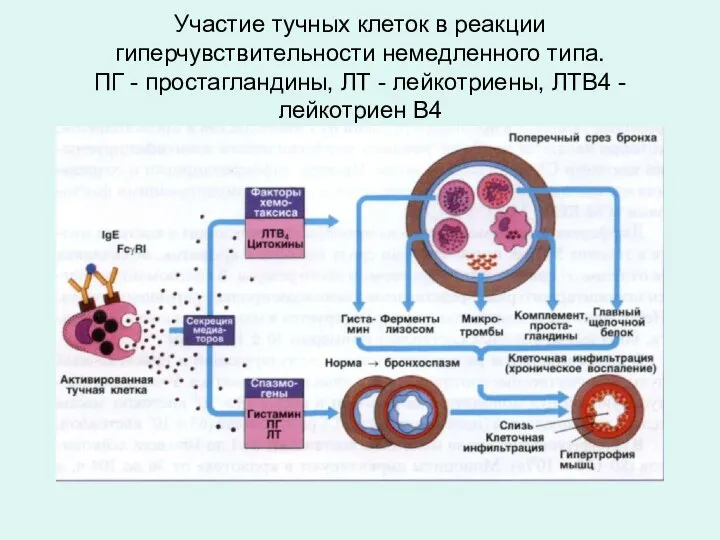
Участие тучных клеток в реакции гиперчувствительности немедленного типа.
ПГ - простагландины, ЛТ
Участие тучных клеток в реакции гиперчувствительности немедленного типа. ПГ - простагландины, ЛТ

Моноцитопоэз. Моноцитарный росток
Монобласт
Промоноцит
Моноцитопоэз. Моноцитарный росток
Монобласт
Промоноцит
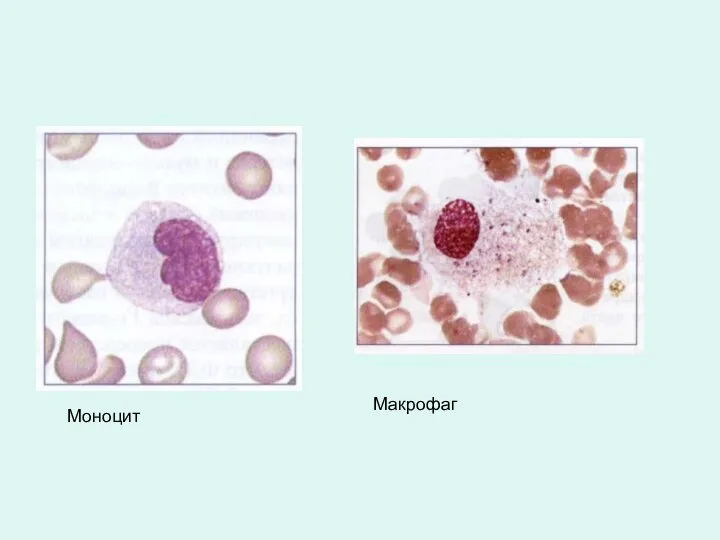
Моноцит
Макрофаг
Моноцит
Макрофаг

Цитохимические маркёры клеток моноцитарно-макрофагальной линии
Неспецифическая эстераза, подавляемая фторидом натрия
Кислая фосфатаза
В процессе
Цитохимические маркёры клеток моноцитарно-макрофагальной линии
Неспецифическая эстераза, подавляемая фторидом натрия
Кислая фосфатаза
В процессе

Мононуклеарные фагоциты
Обновление тканевых макрофагов происходит за счёт притока моноцитов крови
56%
Мононуклеарные фагоциты
Обновление тканевых макрофагов происходит за счёт притока моноцитов крови
56%
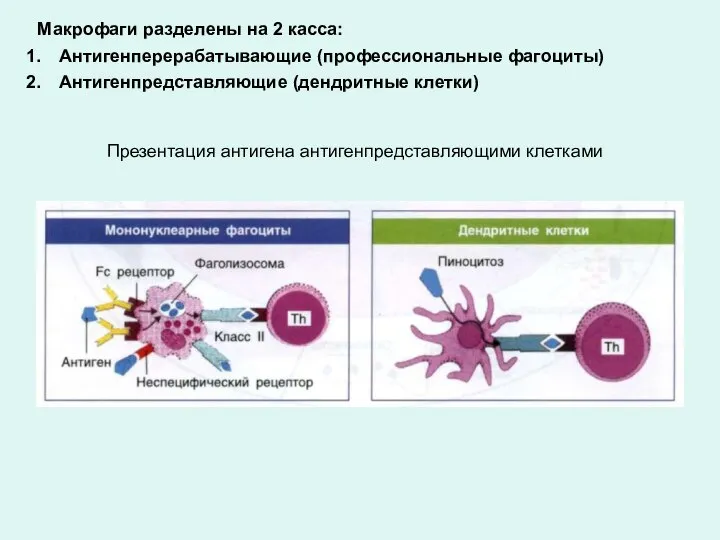
Презентация антигена антигенпредставляющими клетками
Макрофаги разделены на 2 касса:
Антигенперерабатывающие (профессиональные фагоциты)
Антигенпредставляющие
Презентация антигена антигенпредставляющими клетками
Макрофаги разделены на 2 касса:
Антигенперерабатывающие (профессиональные фагоциты)
Антигенпредставляющие

Фолликулярные дендритные клетки
Локализация:
В В-клеточных зонах лимфоузлов
В селезёнке
В мукозо-ассоциированной лимфоидной ткани (MALT)
Представляют
Фолликулярные дендритные клетки
Локализация:
В В-клеточных зонах лимфоузлов
В селезёнке
В мукозо-ассоциированной лимфоидной ткани (MALT)
Представляют

Интердигитальные дендритные клетки
Локализация:
В парокортикальных зонах лимфоузлов
В Т-зависимых зонах селезёнки, миндалин
В мозговом
Интердигитальные дендритные клетки
Локализация:
В парокортикальных зонах лимфоузлов
В Т-зависимых зонах селезёнки, миндалин
В мозговом

Биологическая роль СМФ в процессах обмена веществ
Участие в углеводном обмене (рецепторы
Биологическая роль СМФ в процессах обмена веществ
Участие в углеводном обмене (рецепторы

Секреторные факторы макрофагов
Провоспалительные цитокины (ИЛ-1, ИЛ-6, ИЛ-8, ИЛ-12, ИЛ-18, ФНО-α, интерферон-α,
Секреторные факторы макрофагов
Провоспалительные цитокины (ИЛ-1, ИЛ-6, ИЛ-8, ИЛ-12, ИЛ-18, ФНО-α, интерферон-α,
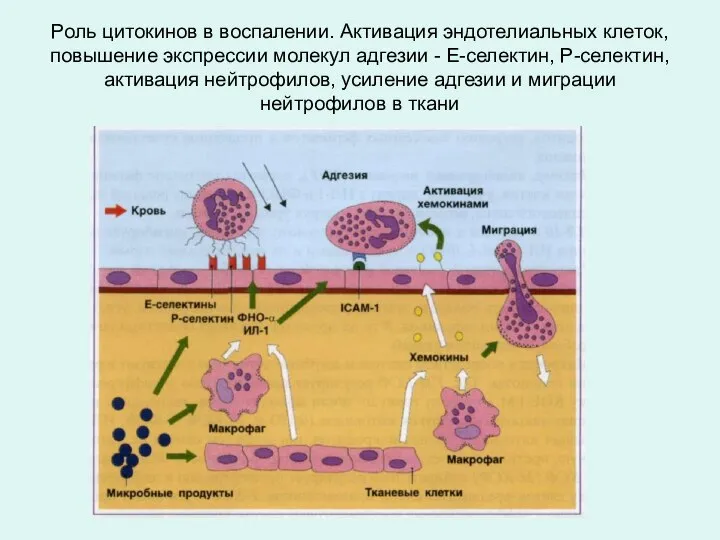
Роль цитокинов в воспалении. Активация эндотелиальных клеток, повышение экспрессии молекул адгезии
Роль цитокинов в воспалении. Активация эндотелиальных клеток, повышение экспрессии молекул адгезии

Лимфоцитопоэз. Лимфоцитарный росток
лимфобласт
пролимфоцит
лимфоцит
Лимфоцитопоэз. Лимфоцитарный росток
лимфобласт
пролимфоцит
лимфоцит
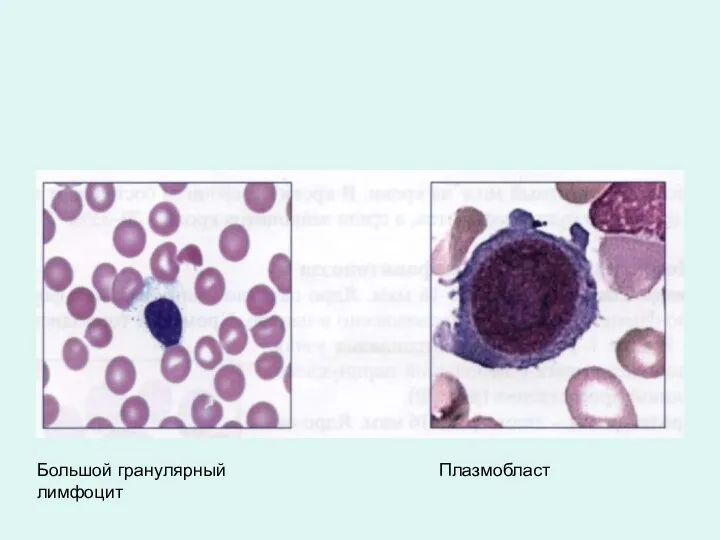
Большой гранулярный лимфоцит
Плазмобласт
Большой гранулярный лимфоцит
Плазмобласт
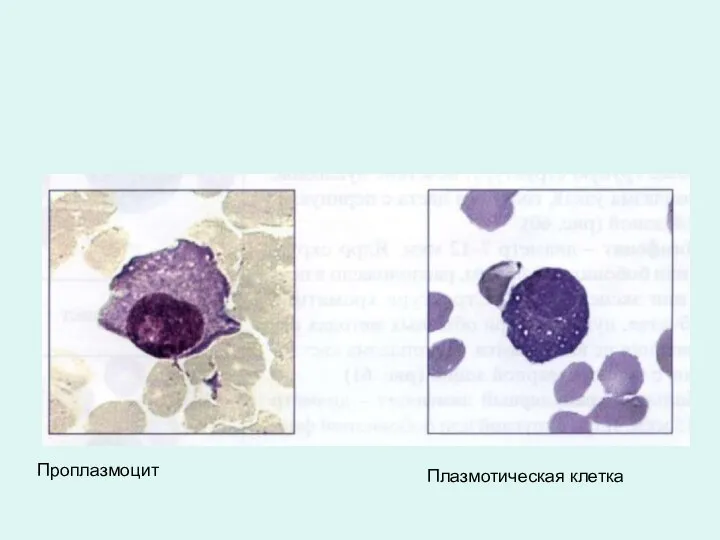
Проплазмоцит
Плазмотическая клетка
Проплазмоцит
Плазмотическая клетка
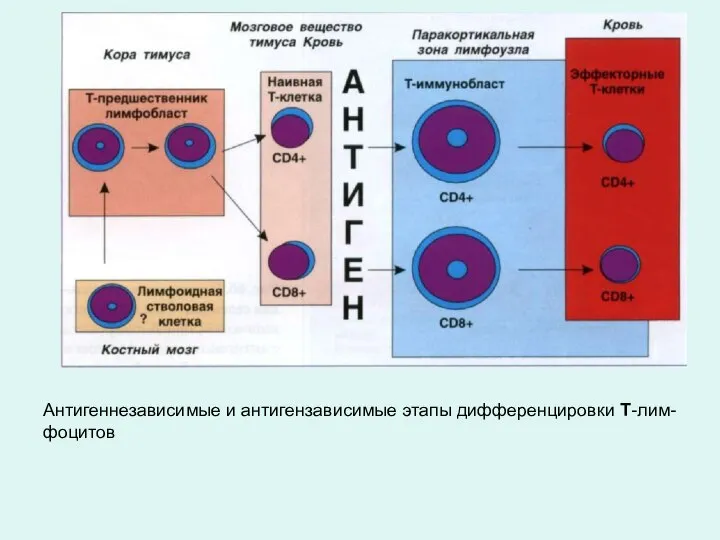
Антигеннезависимые и антигензависимые этапы дифференцировки Т-лим-
фоцитов
Антигеннезависимые и антигензависимые этапы дифференцировки Т-лим-
фоцитов


Дифференцировка В-лимфоцитов
Основной характеристикой В-лимфоцитов является наличие на их мембране рецепторов для
Дифференцировка В-лимфоцитов
Основной характеристикой В-лимфоцитов является наличие на их мембране рецепторов для
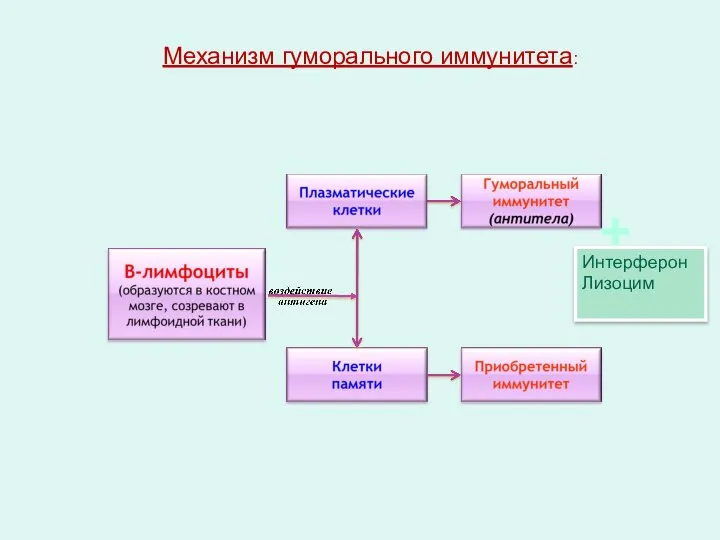
Механизм гуморального иммунитета:
Интерферон
Лизоцим
+
Механизм гуморального иммунитета:
Интерферон
Лизоцим
+

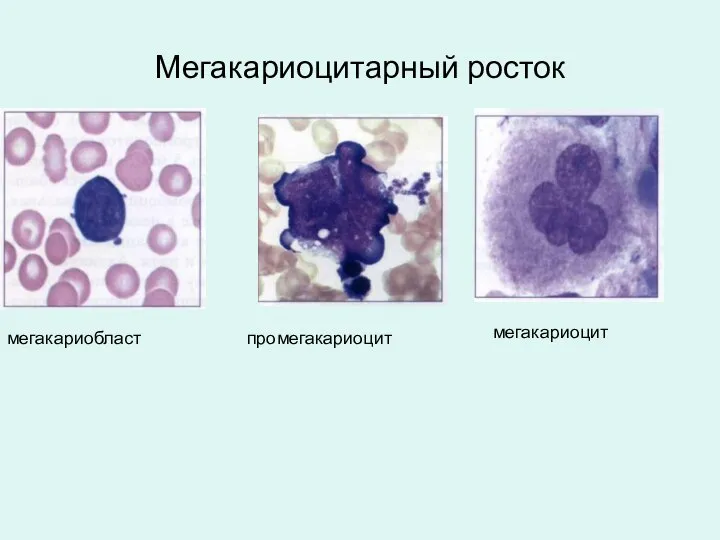
Мегакариоцитарный росток
мегакариобласт
промегакариоцит
мегакариоцит
Мегакариоцитарный росток
мегакариобласт
промегакариоцит
мегакариоцит

Активированные тромбоциты на
поврежденной сосудистой стенке.
При активации происходит округление тромбоцитов
и образование
Активированные тромбоциты на
поврежденной сосудистой стенке.
При активации происходит округление тромбоцитов
и образование
 Доклад об экскурсии в физиотерапевтическое отделение ГУЗ Городская поликлиника 6
Доклад об экскурсии в физиотерапевтическое отделение ГУЗ Городская поликлиника 6 Ситуациялық жағдай
Ситуациялық жағдай Клиническая фармакология противовирусных средств
Клиническая фармакология противовирусных средств Интеллект и интеллектуальная недостаточность
Интеллект и интеллектуальная недостаточность Входящие и выходящие тесты по теме: Контрацепция и аборты
Входящие и выходящие тесты по теме: Контрацепция и аборты Актуальные вопросы лечения ОКИ у детей
Актуальные вопросы лечения ОКИ у детей Лечебные препараты, влияющие на адренергическую иннервацию
Лечебные препараты, влияющие на адренергическую иннервацию Беременность и экстрагенитальные заболевания (заболевания почек, гипертоническая болезнь, сахарный диабет, пороки сердца)
Беременность и экстрагенитальные заболевания (заболевания почек, гипертоническая болезнь, сахарный диабет, пороки сердца) Асептика и антисептика. Организация работы оперблока и хирургического стационара
Асептика и антисептика. Организация работы оперблока и хирургического стационара Лучевая диагностика заболеваний опорно-двигательного аппарата
Лучевая диагностика заболеваний опорно-двигательного аппарата Абдоминальная травма. Исторический обзор
Абдоминальная травма. Исторический обзор Рак почки
Рак почки Морально-этические проблемы контрацепции и стерилизации
Морально-этические проблемы контрацепции и стерилизации Азбука витаминов плюс детские витамины для поддержки зрения. Строение зрительной системы
Азбука витаминов плюс детские витамины для поддержки зрения. Строение зрительной системы Плацентаның жатуы
Плацентаның жатуы Болевой синдром, его виды
Болевой синдром, его виды Pulmonary Nuberculosis
Pulmonary Nuberculosis Асептика. Асептика анықтамасы, міндеттері. Қысқаша тарихи мағлұматтар
Асептика. Асептика анықтамасы, міндеттері. Қысқаша тарихи мағлұматтар Современные аспекты лучевой диагностики лимфопролиферативных заболеваний
Современные аспекты лучевой диагностики лимфопролиферативных заболеваний Сестринская помощь при анемиях и лейкозах
Сестринская помощь при анемиях и лейкозах Половой цикл и его видовые особенности
Половой цикл и его видовые особенности Ойықтарды алдын-алу шаралары
Ойықтарды алдын-алу шаралары Определение остроты зрения
Определение остроты зрения Селективное лазерное плавление SLM/DMP
Селективное лазерное плавление SLM/DMP История развития психопатологии в России
История развития психопатологии в России Нурофен – оригинальный ибупрофен с высокой эффективностью и безопастностью
Нурофен – оригинальный ибупрофен с высокой эффективностью и безопастностью Спинномозговая жидкость, лабораторные методы исследования и их клинико-диагностическое значение
Спинномозговая жидкость, лабораторные методы исследования и их клинико-диагностическое значение Глазное дно
Глазное дно