- Генетика иммуноглобулинов

Содержание
- 2. Список сокращений Ig – иммуноглобулины MHC – главный комплекс гистосовместимости TCR – рецептор Т-лимфоцитов АГ– антиген
- 3. В-ЛИМФОЦИТЫ – продуценты Ig ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
- 4. Функции В-лимфоцитов Противоинфекционная – активность направлена против внеклеточных вирусов и бактерий. Регуляторная – через активность антител
- 5. Компоненты В-системы КОСТНЫЙ МОЗГ лимфоидный орган, обеспечивающий дифференцировку В-лимфоцитов В-КЛЕТКИ В-лимфоциты Антителообразующие клетки Клетки-памяти ИММУНОГЛОБУЛИНЫ молекулы,
- 6. ГУМОРАЛЬНЫЙ ИММУНИТЕТ основные положения В-клетки несут на мембране иммуноглобулины и секретируют их в плазму. Иммуноглобулины (Антитела)
- 7. Весь процесс, приводящий к иммунному ответу организма на вторжение АГ, длится в среднем 10-14 дней. Именно
- 8. ПОЯВЛЕНИЕ И СОЗРЕВАНИЕ В-ЛИМФОЦИТОВ ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
- 9. Схема дифференцировки лимфоцитов
- 11. Этапы «жизни» В-лимфоцита
- 13. Схема дифференцировки В-лимфоцитов без АГ стимула прямое контактное взаимодействие со стромальными элементами – адгезивными молекулами воздействие
- 14. Главное различие между незрелой и иммунокомпетентной В-клеткой - способность синтезировать молекулы иммуноглобулинов
- 15. Схема дифференцировки лимфоцитов с АГ стимуляцией
- 16. ИММУНОГЛОБУЛИНЫ: строение ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
- 17. Функции Ig Иммуноглобулины выполняют в организме позвоночных функцию: гуморальных антител; антиген-связывающих рецепторов В-лимфоцитов.
- 18. структура основной четырехцепочечной единицы иммуноглобулиновых молекул
- 19. Общая схема строения lgG1
- 20. Строение молекулы Ig G Тяжелые (Н) и легкие (L) цепи объединены в единую молекулу с помощью
- 21. Основные фрагменты молекулы Ig
- 22. Взаимодействие антигенсвязывающей области иммуноглобулина с антигеном V-домены легкой и тяжелой цепей (VL и VH) образуют полость
- 23. Принцип АГ-связывающего центра «КЛЮЧ – ЗАМОК»
- 24. Биологические свойства доменов
- 25. ИММУНОГЛОБУЛИНЫ: классы, биологическая роль ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
- 26. Есть две основные формы иммуноглобулинов: Мембрансвязанная (выполняет роль рецептора В-клетки) Секреторная (свободно циркулирующие антитела)
- 27. Классы иммуноглобулинов
- 28. Классы иммуноглобулинов и их биологическая активность
- 29. Функции иммуноглобулинов
- 30. При взаимодействии антигена и Ig- рецептора В-лимфоцита происходит отбор реагирующих В-лимфоцитов.
- 31. Этапы гуморального ответа В-лимфоциты захватывают антиген активируются и процессируют антиген на плазматической мембране в комплексе с
- 32. ИММУНОГЛОБУЛИНЫ: генетическая организация ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
- 33. Реализация генетической информации: классическое представление
- 34. У животных гены Ig и TCR в классическом понимании этого термина отсутствуют. Они возникают de novo
- 35. Локализация зародышевых генов Ig на хромосомах
- 36. Сегментарная организация генов Ig
- 37. Зрелые гены Ig
- 38. Основные варианты зародышевых генов Ig (сегментарная организация) В формировании вариабельных V-доменов Н-цепей участвуют три генных сегмента
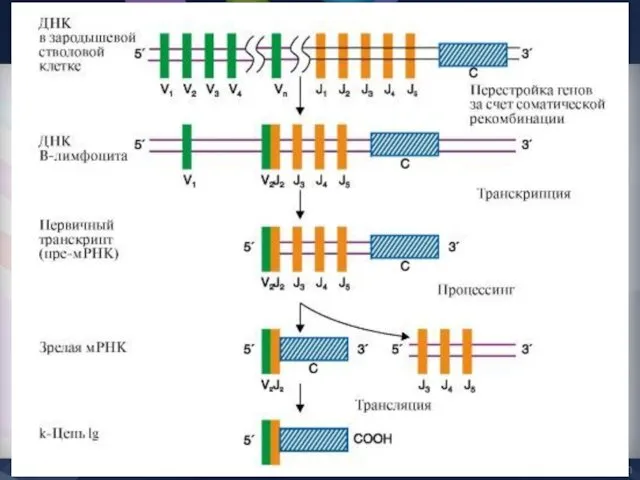
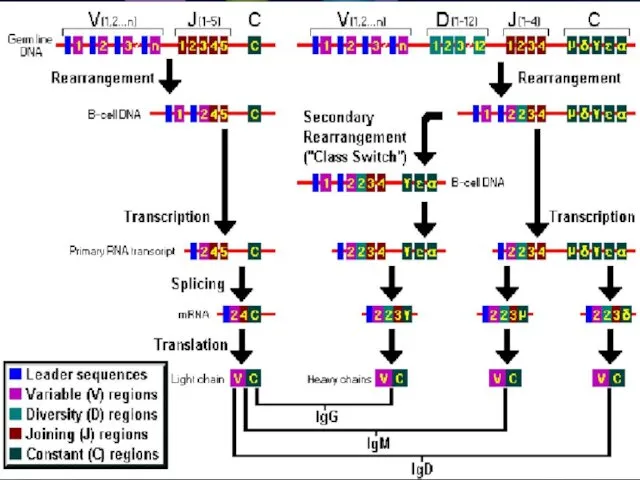
- 39. Программа сборки зрелых генов Ig Перестройки в генах Н-цепей Перестройки генов L-цепей Класс-переключающая перестройка
- 40. Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C) РЕКОМБИНАЦИЯ 1. объединение одного из
- 41. Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C) РЕКОМБИНАЦИЯ 2. объединение одного из
- 42. Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C) РЕКОМБИНАЦИЯ 3. формирование VDJ-Cm-локуса.
- 43. КЛАСС ПЕРЕКЛЮЧАЮЩАЯ ПЕРЕСТРОЙКА Особенность реорганизации генома В-клеток состоит в переключении контроля синтеза Ig одного класса на
- 44. Сегментарная генетическая организация L-цепей Ig æ-локус человека В формировании зрелого гена L-цепи участвуют 3 генных сегмента:
- 45. Этапы перестройки зародышевых генов L цепей 1. перестройка локуса каппа (æ) – æ-локус одного хромосомного гомолога
- 46. Этапы перестройки зародышевых генов L цепей 2. перестройки в лямбда (λ) локусе – начинаются при неудачной
- 47. Этапы перестройки зародышевых генов L цепей 3. апоптоз – если все четыре локуса перестроились неудачно, то
- 50. Класс-переключающая рекомбинация
- 53. Формирование мембранных и секретируемых иммуноглобулинов
- 55. Этапы созревания В-лимфоцитов
- 57. Генерация разнообразия Ig Основной механизм - комбинативное сочетание зародышевых сегментов. Дополнительные: 1. смещение рекомбинационной рамки в
- 58. Схема случайных, разнообразных сочетаний генных сегментов, контролирующих VH-домены тяжелых цепей иммуноглобулинов
- 59. В результате рекомбинации сегментов возможно образование до 30 тыс. вариантов специфических антител.
- 60. С учетом нарушений при рекомбинации, связанных с включением пограничных нуклеотидов справа и слева от D- и
- 62. Скачать презентацию

Список сокращений
Ig – иммуноглобулины
MHC – главный комплекс гистосовместимости
TCR – рецептор
Список сокращений
Ig – иммуноглобулины
MHC – главный комплекс гистосовместимости
TCR – рецептор

В-ЛИМФОЦИТЫ – продуценты Ig
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
В-ЛИМФОЦИТЫ – продуценты Ig
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
Функции В-лимфоцитов
Противоинфекционная – активность направлена против внеклеточных вирусов и бактерий.
Регуляторная
Функции В-лимфоцитов
Противоинфекционная – активность направлена против внеклеточных вирусов и бактерий.
Регуляторная
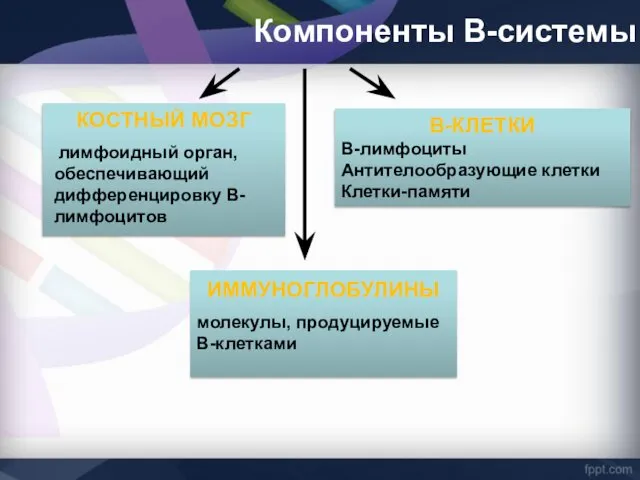
Компоненты В-системы
КОСТНЫЙ МОЗГ
лимфоидный орган, обеспечивающий дифференцировку В-лимфоцитов
В-КЛЕТКИ
В-лимфоциты
Антителообразующие клетки
Клетки-памяти
ИММУНОГЛОБУЛИНЫ
молекулы, продуцируемые В-клетками
Компоненты В-системы
КОСТНЫЙ МОЗГ
лимфоидный орган, обеспечивающий дифференцировку В-лимфоцитов
В-КЛЕТКИ
В-лимфоциты
Антителообразующие клетки
Клетки-памяти
ИММУНОГЛОБУЛИНЫ
молекулы, продуцируемые В-клетками
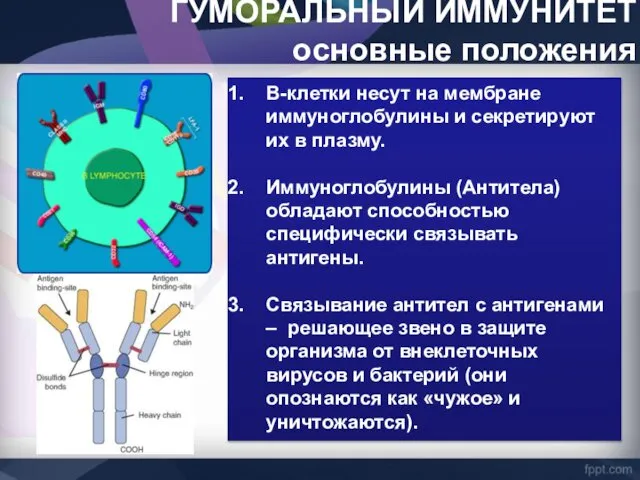
ГУМОРАЛЬНЫЙ ИММУНИТЕТ
основные положения
В-клетки несут на мембране иммуноглобулины и секретируют их в
ГУМОРАЛЬНЫЙ ИММУНИТЕТ
основные положения
В-клетки несут на мембране иммуноглобулины и секретируют их в

Весь процесс, приводящий к иммунному ответу организма на вторжение АГ,
Весь процесс, приводящий к иммунному ответу организма на вторжение АГ,

ПОЯВЛЕНИЕ И СОЗРЕВАНИЕ В-ЛИМФОЦИТОВ
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
ПОЯВЛЕНИЕ И СОЗРЕВАНИЕ В-ЛИМФОЦИТОВ
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
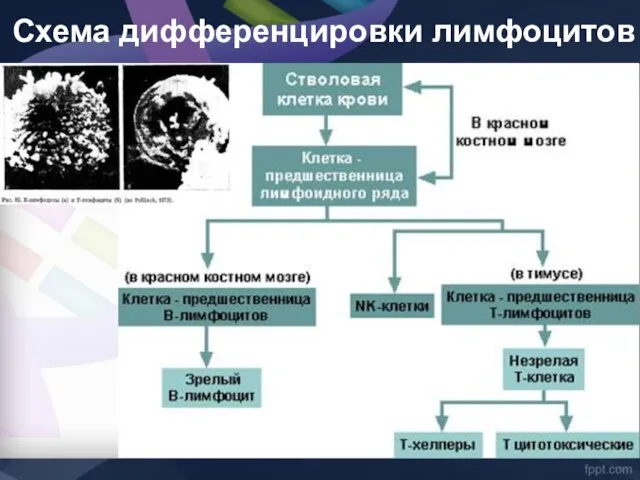
Схема дифференцировки лимфоцитов
Схема дифференцировки лимфоцитов
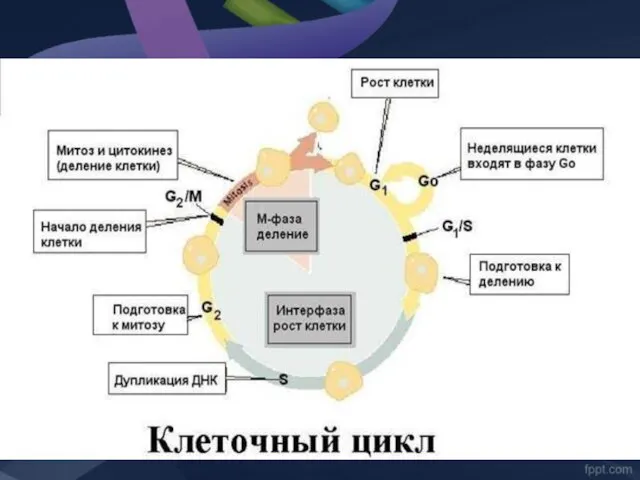
Этапы «жизни» В-лимфоцита
Этапы «жизни» В-лимфоцита

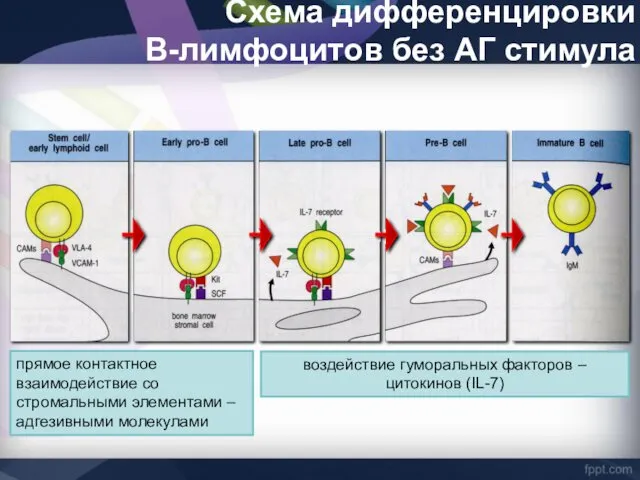
Схема дифференцировки
В-лимфоцитов без АГ стимула
прямое контактное взаимодействие со стромальными элементами
Схема дифференцировки
В-лимфоцитов без АГ стимула
прямое контактное взаимодействие со стромальными элементами

Главное различие между незрелой и иммунокомпетентной В-клеткой - способность синтезировать молекулы
Главное различие между незрелой и иммунокомпетентной В-клеткой - способность синтезировать молекулы
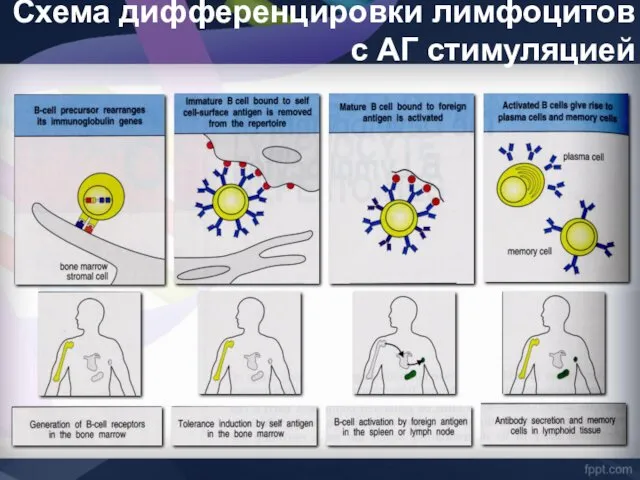
Схема дифференцировки лимфоцитов с АГ стимуляцией
Схема дифференцировки лимфоцитов с АГ стимуляцией

ИММУНОГЛОБУЛИНЫ: строение
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
ИММУНОГЛОБУЛИНЫ: строение
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2

Функции Ig
Иммуноглобулины выполняют в организме позвоночных функцию:
гуморальных антител;
антиген-связывающих рецепторов В-лимфоцитов.
Функции Ig
Иммуноглобулины выполняют в организме позвоночных функцию:
гуморальных антител;
антиген-связывающих рецепторов В-лимфоцитов.
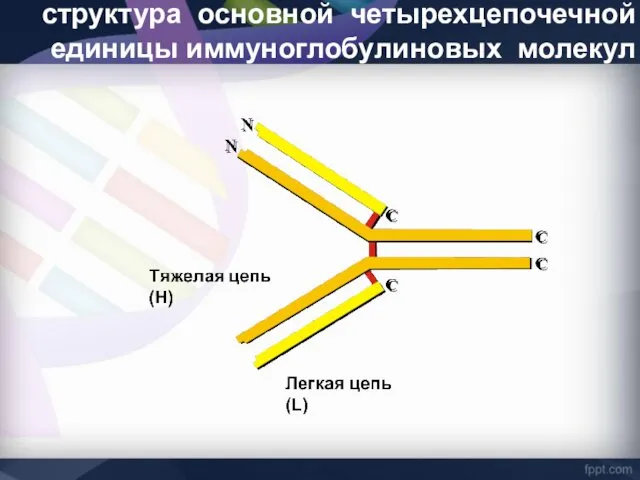
структура основной четырехцепочечной единицы иммуноглобулиновых молекул
структура основной четырехцепочечной единицы иммуноглобулиновых молекул
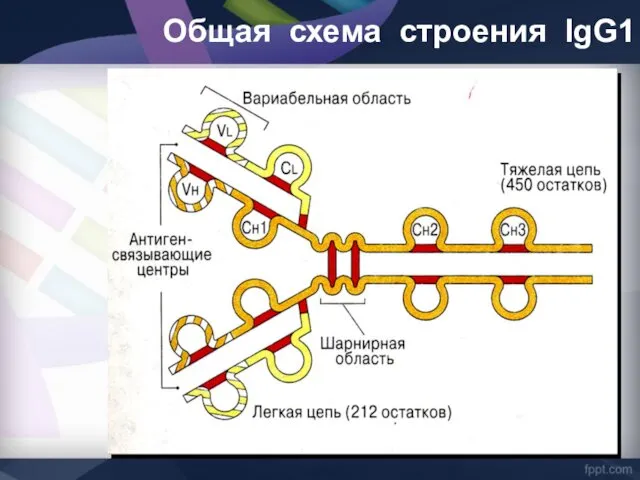
Общая схема строения lgG1
Общая схема строения lgG1
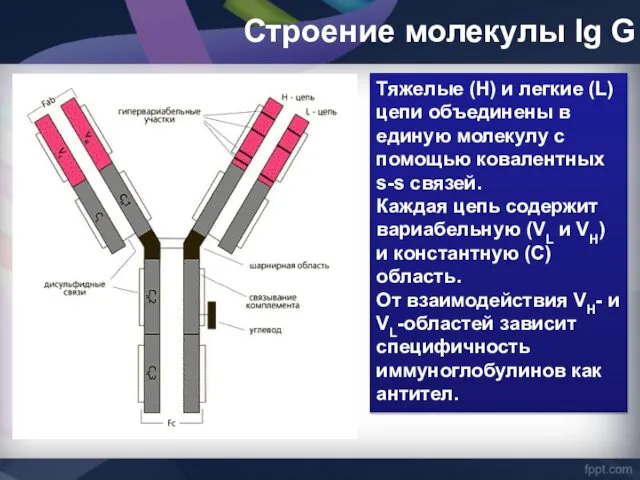
Строение молекулы Ig G
Тяжелые (Н) и легкие (L) цепи объединены в
Строение молекулы Ig G
Тяжелые (Н) и легкие (L) цепи объединены в
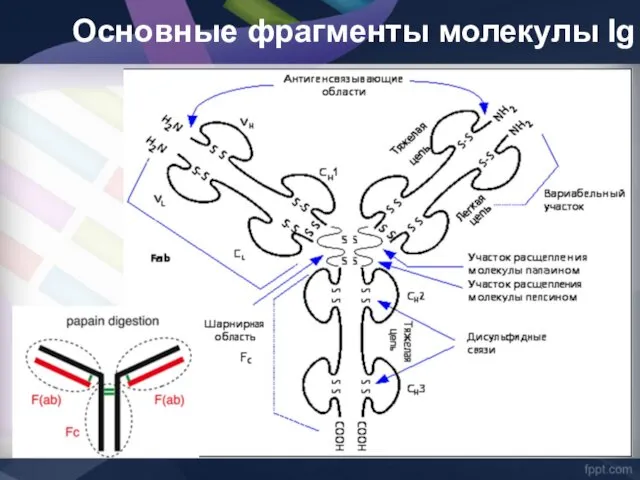
Основные фрагменты молекулы Ig
Основные фрагменты молекулы Ig

Взаимодействие антигенсвязывающей области иммуноглобулина с антигеном
V-домены легкой и тяжелой цепей
Взаимодействие антигенсвязывающей области иммуноглобулина с антигеном
V-домены легкой и тяжелой цепей
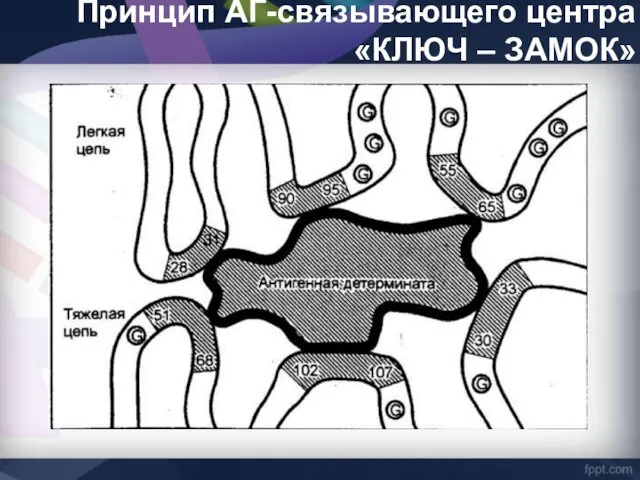
Принцип АГ-связывающего центра «КЛЮЧ – ЗАМОК»
Принцип АГ-связывающего центра «КЛЮЧ – ЗАМОК»
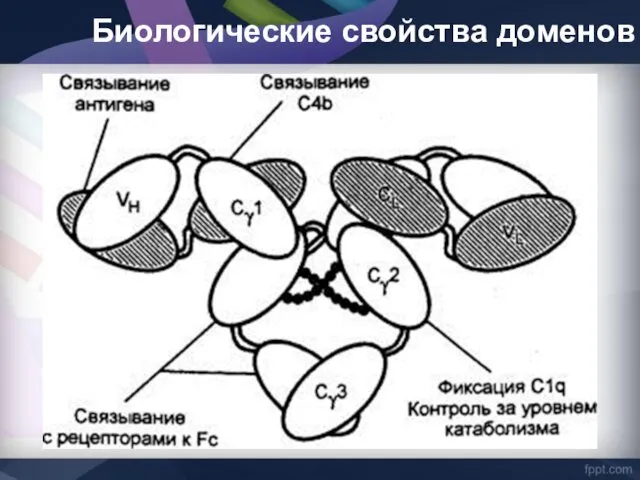
Биологические свойства доменов
Биологические свойства доменов

ИММУНОГЛОБУЛИНЫ: классы, биологическая роль
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
ИММУНОГЛОБУЛИНЫ: классы, биологическая роль
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2

Есть две основные формы иммуноглобулинов:
Мембрансвязанная (выполняет роль рецептора В-клетки)
Секреторная (свободно
Есть две основные формы иммуноглобулинов:
Мембрансвязанная (выполняет роль рецептора В-клетки)
Секреторная (свободно
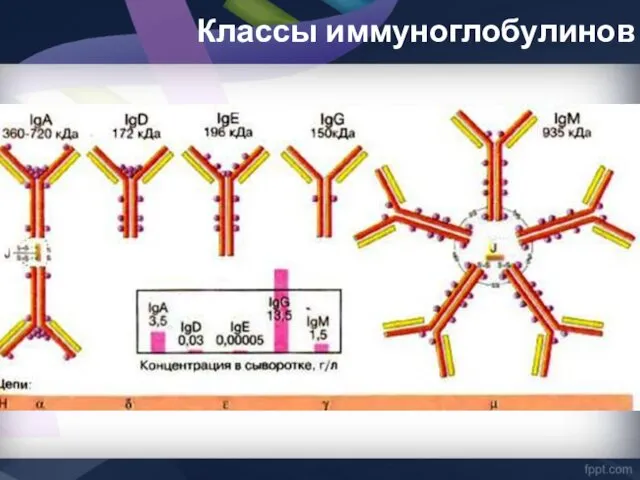
Классы иммуноглобулинов
Классы иммуноглобулинов
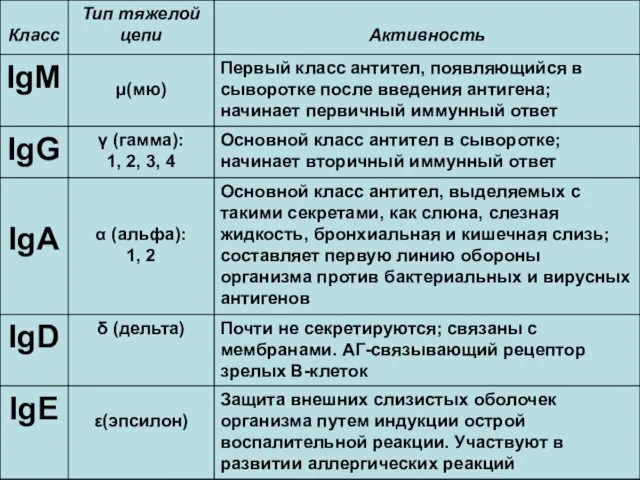
Классы иммуноглобулинов и их биологическая активность
Классы иммуноглобулинов и их биологическая активность

Функции иммуноглобулинов
Функции иммуноглобулинов
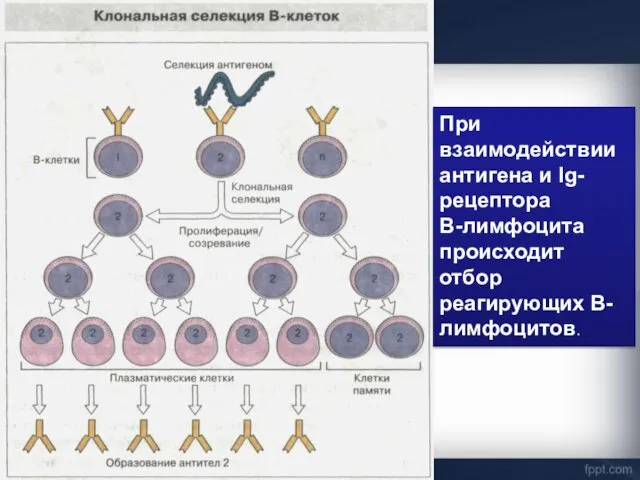
При взаимодействии антигена и Ig- рецептора
В-лимфоцита происходит отбор реагирующих В-лимфоцитов.
При взаимодействии антигена и Ig- рецептора
В-лимфоцита происходит отбор реагирующих В-лимфоцитов.

Этапы гуморального ответа
В-лимфоциты захватывают антиген активируются и процессируют антиген на плазматической
Этапы гуморального ответа
В-лимфоциты захватывают антиген активируются и процессируют антиген на плазматической

ИММУНОГЛОБУЛИНЫ: генетическая организация
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
ИММУНОГЛОБУЛИНЫ: генетическая организация
ГЕНЕТИКА ИММУНИТЕТА. Лекция 2
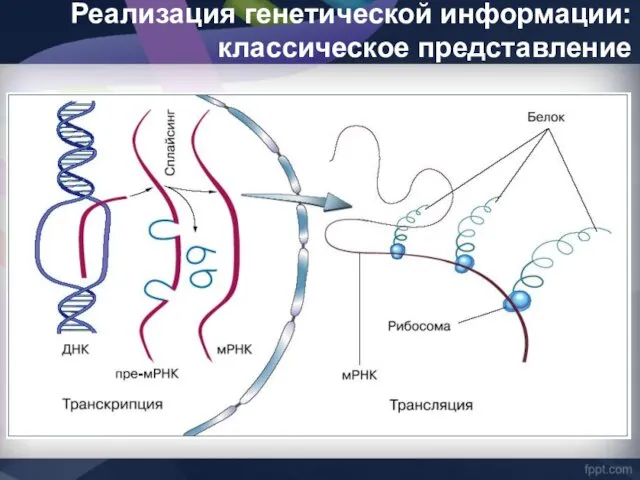
Реализация генетической информации: классическое представление
Реализация генетической информации: классическое представление

У животных гены Ig и TCR в классическом понимании этого термина
У животных гены Ig и TCR в классическом понимании этого термина
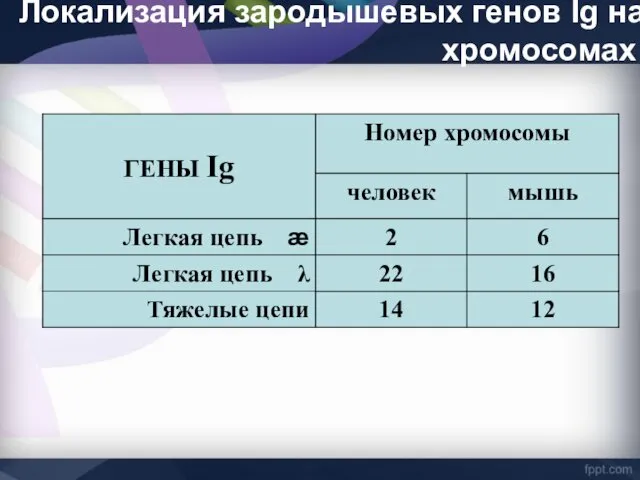
Локализация зародышевых генов Ig на хромосомах
Локализация зародышевых генов Ig на хромосомах
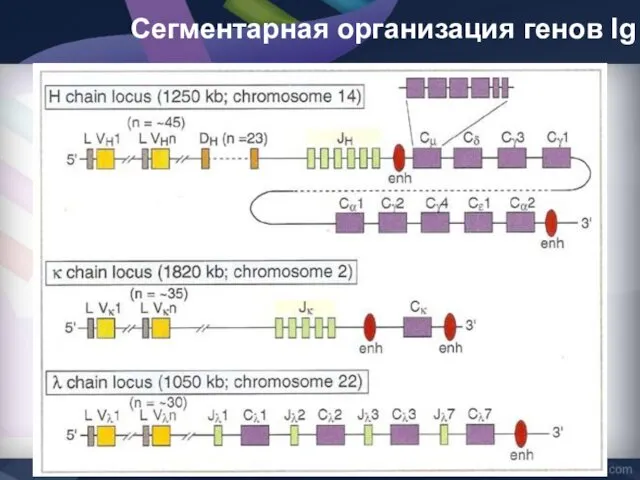
Сегментарная организация генов Ig
Сегментарная организация генов Ig
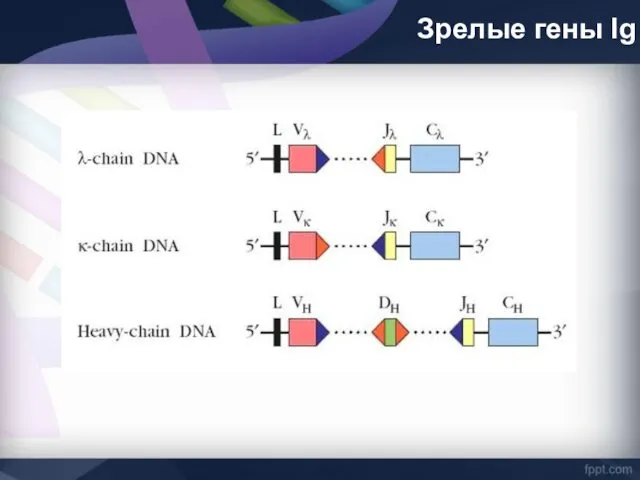
Зрелые гены Ig
Зрелые гены Ig
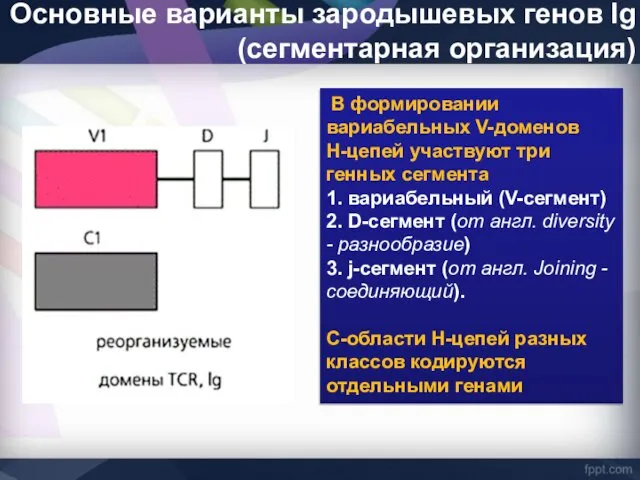
Основные варианты зародышевых генов Ig (сегментарная организация)
В формировании вариабельных V-доменов
Основные варианты зародышевых генов Ig (сегментарная организация)
В формировании вариабельных V-доменов

Программа сборки зрелых генов Ig
Перестройки в генах Н-цепей
Перестройки генов L-цепей
Класс-переключающая перестройка
Программа сборки зрелых генов Ig
Перестройки в генах Н-цепей
Перестройки генов L-цепей
Класс-переключающая перестройка
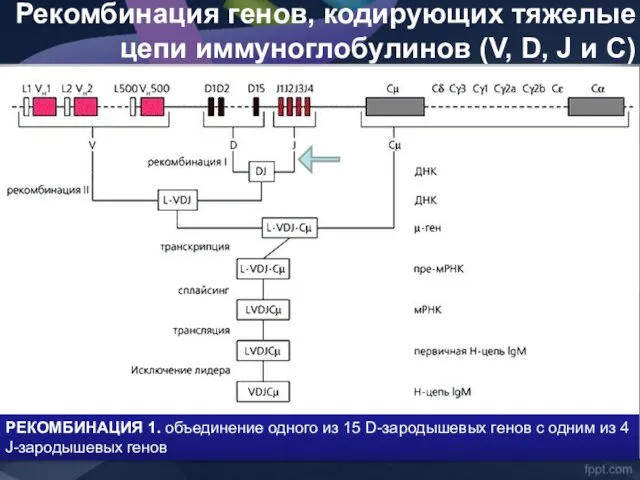
Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C)
Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C)
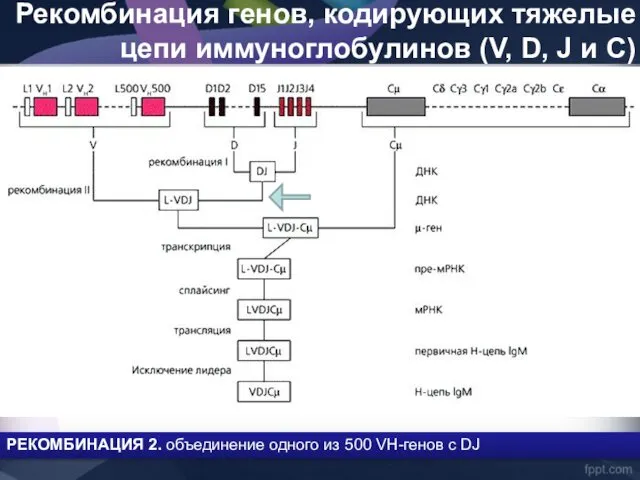
Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C)
Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C)
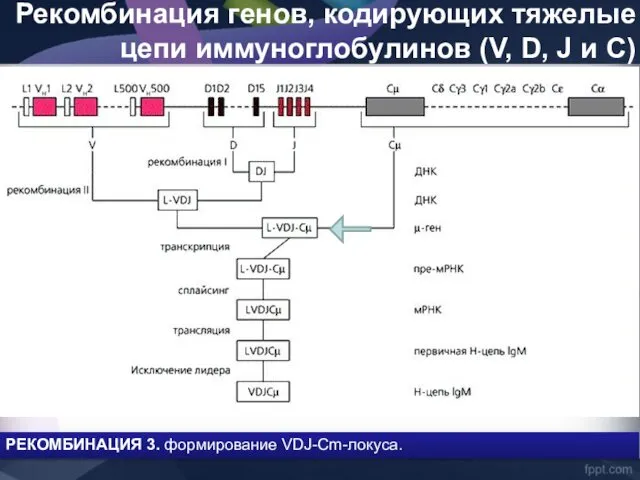
Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C)
Рекомбинация генов, кодирующих тяжелые цепи иммуноглобулинов (V, D, J и C)
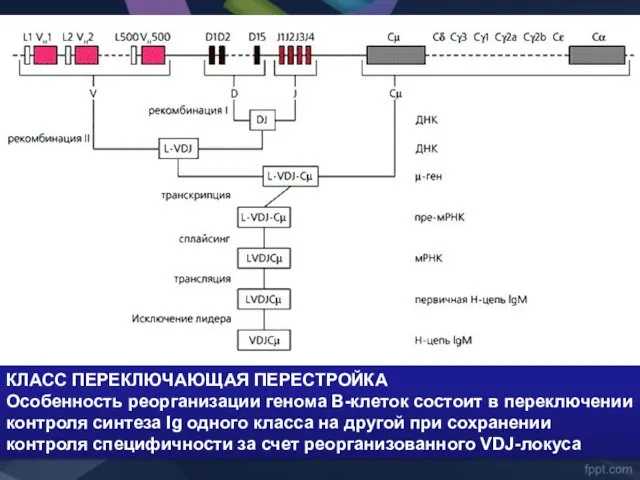
КЛАСС ПЕРЕКЛЮЧАЮЩАЯ ПЕРЕСТРОЙКА
Особенность реорганизации генома В-клеток состоит в переключении контроля синтеза
КЛАСС ПЕРЕКЛЮЧАЮЩАЯ ПЕРЕСТРОЙКА
Особенность реорганизации генома В-клеток состоит в переключении контроля синтеза

Сегментарная генетическая организация L-цепей Ig
æ-локус человека
В формировании зрелого гена L-цепи участвуют
Сегментарная генетическая организация L-цепей Ig
æ-локус человека
В формировании зрелого гена L-цепи участвуют
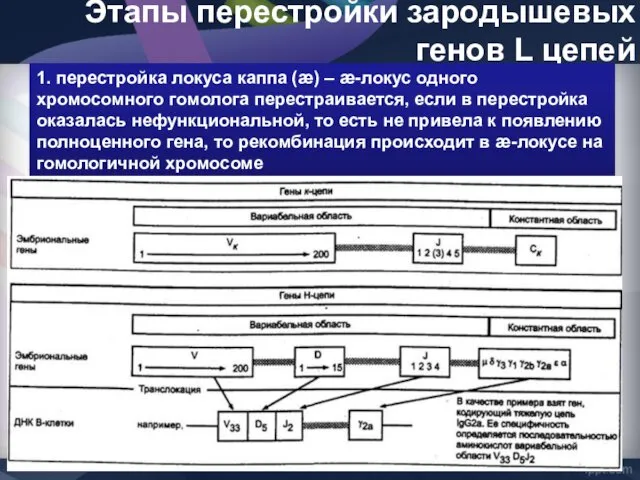
Этапы перестройки зародышевых генов L цепей
1. перестройка локуса каппа (æ) –
Этапы перестройки зародышевых генов L цепей
1. перестройка локуса каппа (æ) –
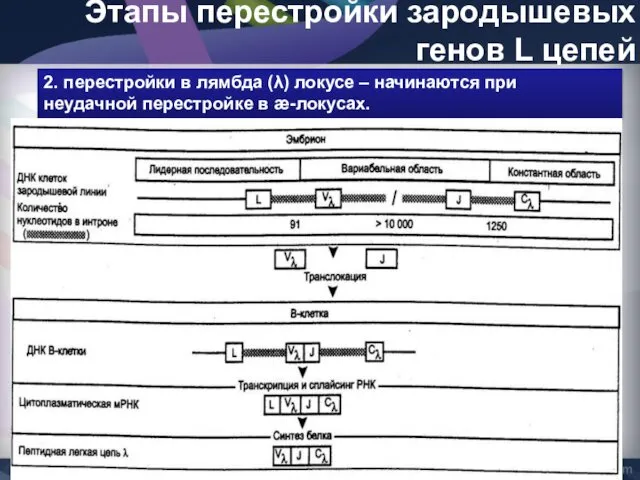
Этапы перестройки зародышевых генов L цепей
2. перестройки в лямбда (λ) локусе
Этапы перестройки зародышевых генов L цепей
2. перестройки в лямбда (λ) локусе

Этапы перестройки зародышевых генов L цепей
3. апоптоз – если все четыре
Этапы перестройки зародышевых генов L цепей
3. апоптоз – если все четыре

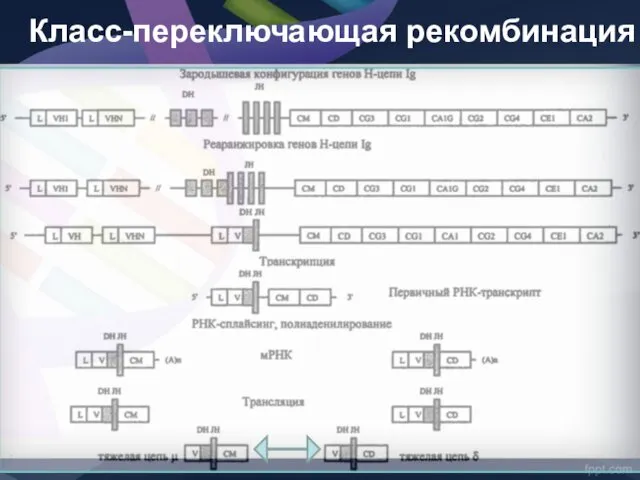
Класс-переключающая рекомбинация
Класс-переключающая рекомбинация

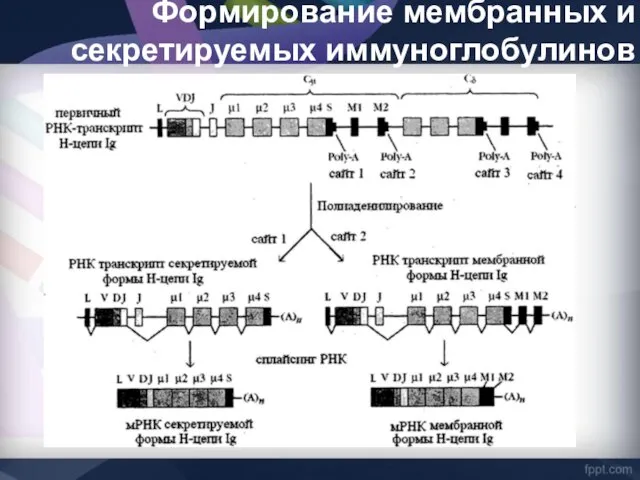
Формирование мембранных и секретируемых иммуноглобулинов
Формирование мембранных и секретируемых иммуноглобулинов


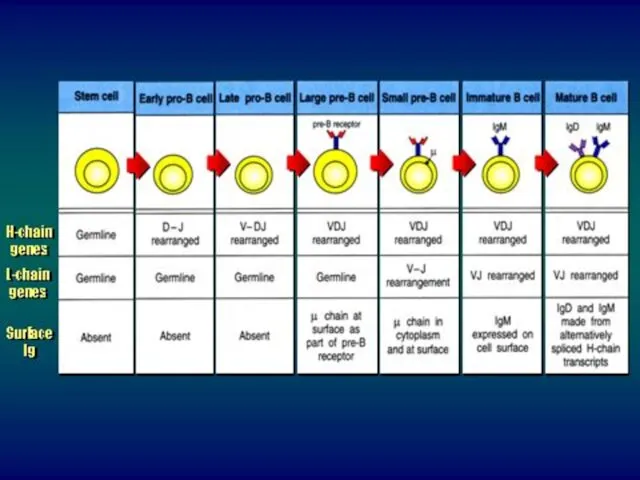
Этапы созревания В-лимфоцитов
Этапы созревания В-лимфоцитов

Генерация разнообразия Ig
Основной механизм - комбинативное сочетание зародышевых сегментов.
Дополнительные:
1. смещение
Генерация разнообразия Ig
Основной механизм - комбинативное сочетание зародышевых сегментов.
Дополнительные:
1. смещение

Схема случайных, разнообразных сочетаний генных сегментов, контролирующих VH-домены тяжелых цепей иммуноглобулинов
Схема случайных, разнообразных сочетаний генных сегментов, контролирующих VH-домены тяжелых цепей иммуноглобулинов

В результате рекомбинации сегментов возможно образование до 30 тыс. вариантов специфических антител.
В результате рекомбинации сегментов возможно образование до 30 тыс. вариантов специфических антител.

С учетом нарушений при рекомбинации, связанных с включением пограничных нуклеотидов справа
С учетом нарушений при рекомбинации, связанных с включением пограничных нуклеотидов справа
 Сестринская помощь при повреждениях мягких тканей, костей конечностей
Сестринская помощь при повреждениях мягких тканей, костей конечностей Сообщение печальных новостей
Сообщение печальных новостей Угревая болезнь
Угревая болезнь Пять эффективных способов решения конфликтов
Пять эффективных способов решения конфликтов Эпилепсия. Клинические проявления
Эпилепсия. Клинические проявления Механизм образования билирубина
Механизм образования билирубина Блокаторы калиевых каналов. Амиодарон
Блокаторы калиевых каналов. Амиодарон Паркинсон ауруы
Паркинсон ауруы Зейін және ес бұзылыстарының клиникалық сипаттамасы
Зейін және ес бұзылыстарының клиникалық сипаттамасы Сестринский персонал в программах профилактики ВИЧ. Лекция 1
Сестринский персонал в программах профилактики ВИЧ. Лекция 1 Бронх демікпесі
Бронх демікпесі Выделительная система
Выделительная система Балалардағы қант диабетінің алдын алу
Балалардағы қант диабетінің алдын алу Дәрігер-науқас-мейірбеке қарым-қатнасы
Дәрігер-науқас-мейірбеке қарым-қатнасы Бешенство (Rabies)
Бешенство (Rabies) Пункция заднего свода влагалища (кульдоцентез)
Пункция заднего свода влагалища (кульдоцентез) ЭКГ при нарушениях ритма и проводимости
ЭКГ при нарушениях ритма и проводимости Предпрофильная подготовка. Тип темперамента
Предпрофильная подготовка. Тип темперамента Формирование благоприятного социально-психологического климата в классном коллективе учащихся и родителей
Формирование благоприятного социально-психологического климата в классном коллективе учащихся и родителей Трахеостомия
Трахеостомия Влагалищные пессарии: виды, показания, правила подбора, плюсы и минусы использования
Влагалищные пессарии: виды, показания, правила подбора, плюсы и минусы использования Школа Флоренс Найнтингейл
Школа Флоренс Найнтингейл Алғашқы медико-санитарлық көмекті ұйымдастыру
Алғашқы медико-санитарлық көмекті ұйымдастыру Артериальная гипертензия и гипотензия во время анестезии и ближайшем послеоперационном периоде
Артериальная гипертензия и гипотензия во время анестезии и ближайшем послеоперационном периоде Осанка. Комплекс упражнений для формирования правильной осанки
Осанка. Комплекс упражнений для формирования правильной осанки Балалардың асқазан - ішек аурулары туралы түсінік беру
Балалардың асқазан - ішек аурулары туралы түсінік беру Гипреактивные дети
Гипреактивные дети Дифференциальная диагностика. Миастения
Дифференциальная диагностика. Миастения