- Молекулярно-генетическая диагностика. ПЦР и его модификации. Курс 3 ЦИОП Медицина будущего

Содержание
- 2. Молекулярно-генетическая диагностика Основные стадии молекулярно-генетического анализа: Выделение ДНК или РНК из клинического материала Амплификация анализируемых участков
- 3. Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских исследований? Клеточные «суспензии» Ликвор, плазма
- 4. Метод сорбции нуклеиновых кислот на силиконовом носителе В основе лежит избирательная сорбция НК в присутствии хаотропной
- 5. Метод фенол-хлороформной экстракции Принцип метода основан на том, что НК является полярной молекулой и не растворяется
- 6. Определение количества и качества нуклеиновых кислот Качественный метод – электрофорез в агарозном геле нуклеиновой кислоты и
- 7. ПЦР – репликация ДНК in vitro В 1993 г. Kary B. Mullis стал лауреатом Нобелевской премии
- 8. Метод ПЦР – краткое напоминание
- 9. Компоненты ПЦР Буфер (Tris-HCl, pH 8.0-8.8, KCl) MgCl2 (1-3 mM) Праймеры (0.4 мкМоль каждого) dNTP (40-200
- 10. Ферменты ПЦР
- 11. Праймеры для ПЦР
- 13. Оптимизация ПЦР
- 14. Оптимизация ПЦР
- 15. Вещества, улучшающие специфичность и/или выход ПЦР
- 16. Другие вещества, влияющие на ПЦР
- 17. Влияние красителей на на ПЦР
- 18. ПЦР: правила хорошего тона
- 19. Программируемые термоциклеры Что важно учитывать при выборе: Формат (стандартные 0,2 мл, или 0,1 мл, или 0,5
- 20. Способы визуализации ПЦР «по конечной точке» Детекция по конечной точке: FLASH Электрофорез в геле
- 21. Электрофорез в агарозном геле Гель формируется за счет нековалентных связей. Имеет относительно низкую разрешающую способность. Раньше
- 23. Электрофорез в полиакриламидном геле Большая разрешающая способность. Часто используют в молекулярно-генетической диагностике. Красить лучше нитратом серебра
- 24. Гетеродуплексный анализ Метод основан на том, что при ренатурации смеси нормальных и мутантных аллелей наряду с
- 25. SSCP-анализ SSCP – Single Strand Conformation Polymorphism. Метод основан на разной подвижности при электрофорезе в ПААГ
- 26. Мультиплексная ПЦР При увеличении количества локусов в мультиплексной ПЦР, практически, экспоненциально растут затраты времени на отработку
- 27. ПЦР-ПДРФ ПДРФ – полиморфизм длин рестрикционных фрагментов. ПЦР-продукт обрабатывают рестриктазой, которая гидролизует только один из аллелей
- 28. Аллель-специфичная ПЦР Основная проблема АС-ПЦР – это неспецифический синтез с частично комплементарного праймера. Решения: вводить неспаренные
- 29. Метилчувствительная ПЦР Схема эксперимента Преимущества: Дешевизна и доступность реактивов и расходных материалов Высокая чувствительность Недостатки: Можно
- 30. Преимущества: Возможность анализировать CpG независимо от локализации Стандартизированная методика ПЦР-продукт можно секвенировать и определить статус метилирования
- 31. Фрагментный анализ STR-маркеров на капиллярном секвенаторе 1. Сбор исходных данных о флуоресценции в режиме реального времени
- 32. Секвенирование ПЦР-продуктов по Сэнгеру
- 33. Гнездовая (nested) ПЦР
- 34. 1 - двухцепочечная ДНК, с экзонами и интронами. Праймер 1 (прямой) расположен в 1 экзоне, праймер
- 35. Hot-Start ПЦР
- 36. Требования к организации работы ПЦР-лабораторий Если есть реал-тайм ПЦР, то зона Г не нужна. Д –
- 37. ПЦР – это качественный или количественный анализ? Если оценивать по конечной точке - качественный Nn =
- 38. ПЦР – это качественный или количественный анализ? (продолжение) Если измерять концентрации продуктов в log-фазе - количественный
- 39. Количественный анализ – ПЦР в реальном времени Сигнал от флуоресцентной метки, пропорциональный количеству ПЦР-продукта, считывают на
- 40. Детектирующие амплификаторы ДТ-96 (ДНК-технология, Россия): 4 канала, 96-луночная плашка Rotorgene 6000 (Corbett Research, Австралия): нельзя использовать
- 41. ПЦР в реальном времени (ПЦР-РВ) Детектирующий амплификатор определяет количество ПЦР-продукта на каждом цикле, но не молекулы
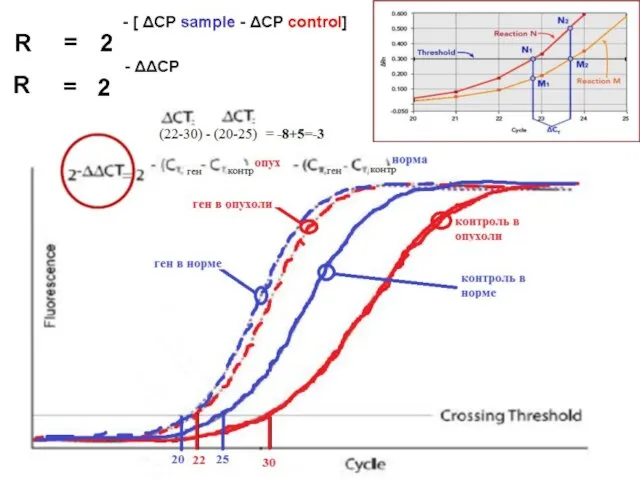
- 42. Пороговый метод сравнения графиков накопления ДНК Суть порогового метода заключается в том, чтобы определить момент Ct,
- 43. Пороговый метод сравнения графиков накопления ДНК Сначала необходимо вычислить Е и произвести нормировку α. Для нормировки
- 44. Варианты ПЦР в реальном времени Системы детекции Специфичные Неспецифичные Интеркалирующие красители Амплифлюры Скорпионы Свободные меченые зонды
- 45. Недостатки: 1. взаимодействует с неспецифическими продуктами амплификации 2. взаимодействует с димерами праймеров 3. невозможно использовать в
- 46. Амплифлюры
- 47. Зонды TaqMan
- 48. Молекулярный маяк представляет собой меченый красителем олигонуклеотид (25-40 нуклеотидов), который образует шпилечную структуру со стеблем и
- 49. Флуорофоры и гасители ПЦР-РВ Флуорофоры и гасители флуоресценции. Как правило, корректное проведение ПЦР требует постановки нескольких
- 50. Определение эффективности ПЦР-РВ Вычисление Е по последовательным разведениям образца (измерение абсолютного количества ДНК на определенном цикле)
- 51. Определение эффективности ПЦР-РВ Вычисление Е по последовательным разведениям образца Вычисление Е по углу наклона прямого участка
- 52. Абсолютное и относительное определение количества исходной ДНК Формула N0 = N0rE(Ctr – Ct) необходима тогда, когда
- 53. Абсолютное и относительное определение количества исходной ДНК (продолжение) Предполагая, что условия ПЦР-РВ оптимизированы и эффективности реакции
- 54. Количественное определение вирусной нагрузки Этапы эксперимента: Пробоподготовка (выделение вирусной РНК из плазмы крови) Обратная транскрипция (получение
- 55. ДНК-диагностика онкогематологических заболеваний Полная ремиссия: отсутствие клинических симптомов болезни (клиническая ремиссия) и присутствие в миелограмме менее
- 56. Метилспецифическая ПЦР-РВ при изучении генов-супрессоров Метилирование – это присоединение метильной группы в позиции N5 пиримидинового кольца
- 57. Что надо сделать до проведения ПЦР в реальном времени для изучения экспрессии гена? Метод кисло-фенольной экстракции
- 58. Выбор референсных генов Референсные гены, расположенные в порядке увеличения стабильности их экспрессии (Vandesompele et al., 2002).
- 60. Цифровая ПЦР (Digital PCR) Схема метода по сравнению экспрессии генов: Выделение РНК, обратная транскрипция, разведение препарата
- 61. Выходящий тестовый контроль 1. Увеличение количества ПЦР-продуктов при эффективности ПЦР, равной 2, происходит: линейно логарифмически стохастически
- 64. Скачать презентацию

Молекулярно-генетическая диагностика
Основные стадии молекулярно-генетического анализа:
Выделение ДНК или РНК из клинического материала
Амплификация
Молекулярно-генетическая диагностика
Основные стадии молекулярно-генетического анализа:
Выделение ДНК или РНК из клинического материала
Амплификация

Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских
Из какого материала приходится выделять ДНК и РНК при проведении биомедицинских
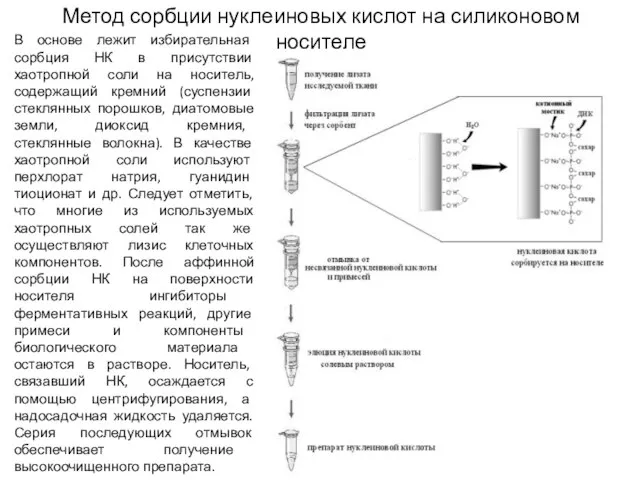
Метод сорбции нуклеиновых кислот на силиконовом носителе
В основе лежит избирательная сорбция
Метод сорбции нуклеиновых кислот на силиконовом носителе
В основе лежит избирательная сорбция
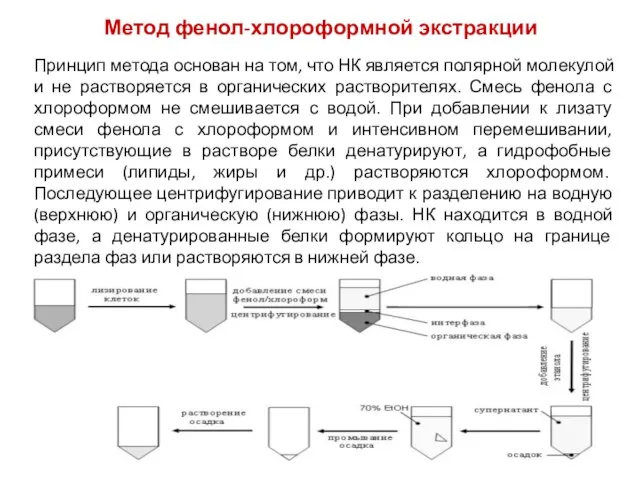
Метод фенол-хлороформной экстракции
Принцип метода основан на том, что НК является полярной
Метод фенол-хлороформной экстракции
Принцип метода основан на том, что НК является полярной
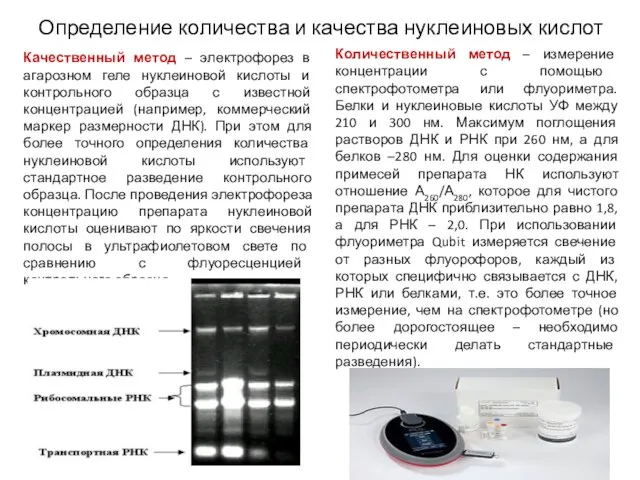
Определение количества и качества нуклеиновых кислот
Качественный метод – электрофорез в агарозном
Определение количества и качества нуклеиновых кислот
Качественный метод – электрофорез в агарозном

ПЦР – репликация ДНК in vitro
В 1993 г. Kary B. Mullis
ПЦР – репликация ДНК in vitro
В 1993 г. Kary B. Mullis

Метод ПЦР – краткое напоминание
Метод ПЦР – краткое напоминание

Компоненты ПЦР
Буфер (Tris-HCl, pH 8.0-8.8, KCl)
MgCl2 (1-3 mM)
Праймеры (0.4 мкМоль каждого)
dNTP
Компоненты ПЦР
Буфер (Tris-HCl, pH 8.0-8.8, KCl)
MgCl2 (1-3 mM)
Праймеры (0.4 мкМоль каждого)
dNTP
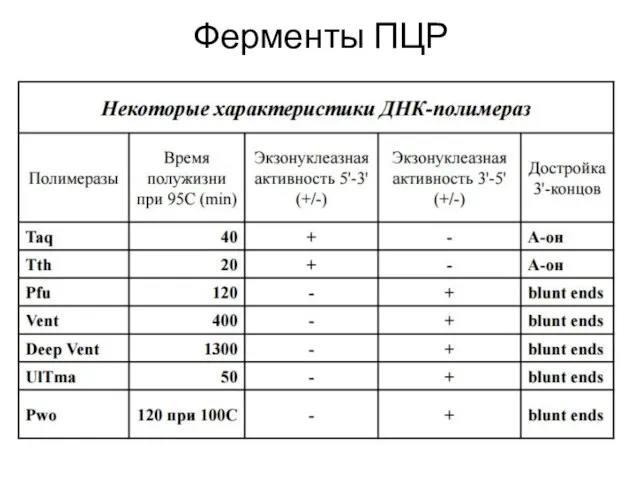
Ферменты ПЦР
Ферменты ПЦР

Праймеры для ПЦР
Праймеры для ПЦР


Оптимизация ПЦР
Оптимизация ПЦР
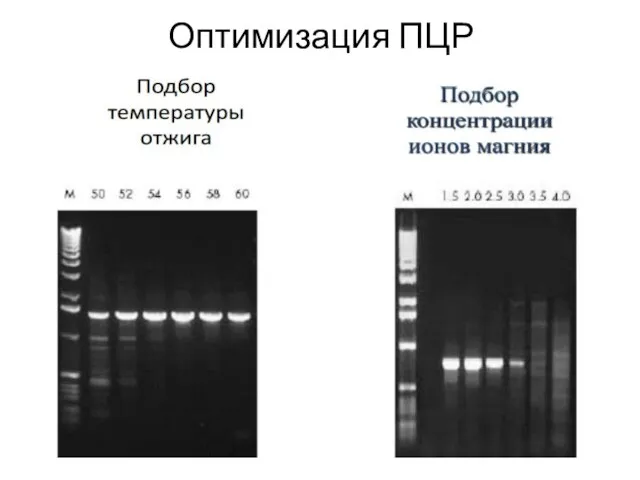
Оптимизация ПЦР
Оптимизация ПЦР

Вещества, улучшающие специфичность и/или выход ПЦР
Вещества, улучшающие специфичность и/или выход ПЦР

Другие вещества, влияющие на ПЦР
Другие вещества, влияющие на ПЦР

Влияние красителей на на ПЦР
Влияние красителей на на ПЦР

ПЦР: правила хорошего тона
ПЦР: правила хорошего тона

Программируемые термоциклеры
Что важно учитывать при выборе:
Формат (стандартные 0,2 мл, или 0,1
Программируемые термоциклеры
Что важно учитывать при выборе:
Формат (стандартные 0,2 мл, или 0,1
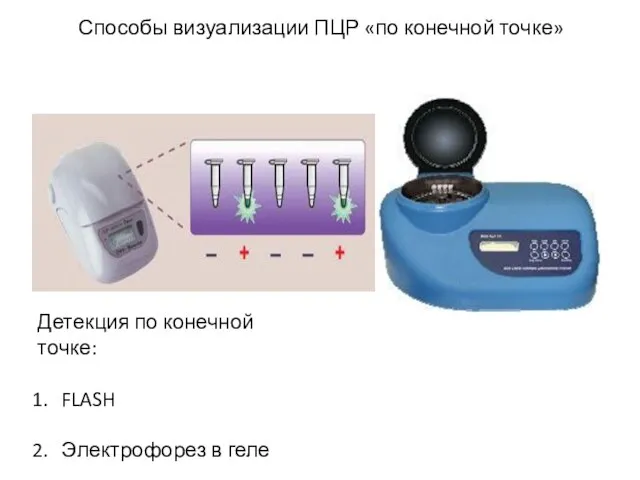
Способы визуализации ПЦР «по конечной точке»
Детекция по конечной точке:
FLASH
Электрофорез в геле
Способы визуализации ПЦР «по конечной точке»
Детекция по конечной точке:
FLASH
Электрофорез в геле
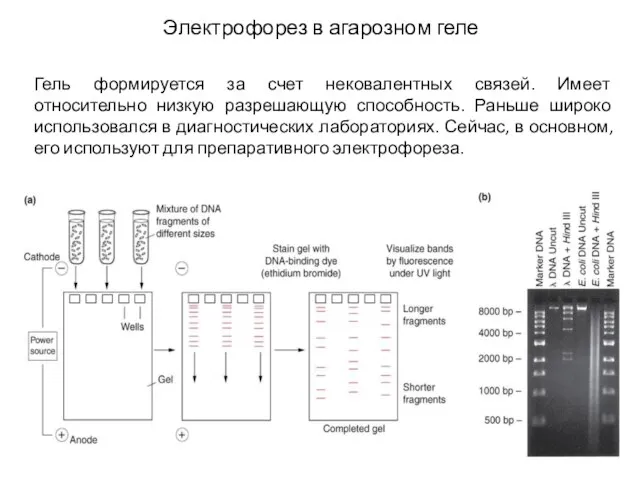
Электрофорез в агарозном геле
Гель формируется за счет нековалентных связей. Имеет относительно
Электрофорез в агарозном геле
Гель формируется за счет нековалентных связей. Имеет относительно
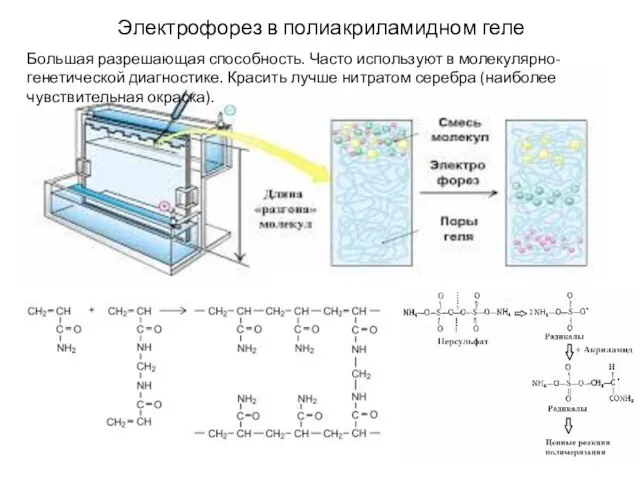
Электрофорез в полиакриламидном геле
Большая разрешающая способность. Часто используют в молекулярно-генетической диагностике.
Электрофорез в полиакриламидном геле
Большая разрешающая способность. Часто используют в молекулярно-генетической диагностике.

Гетеродуплексный анализ
Метод основан на том, что при ренатурации смеси нормальных и
Гетеродуплексный анализ
Метод основан на том, что при ренатурации смеси нормальных и
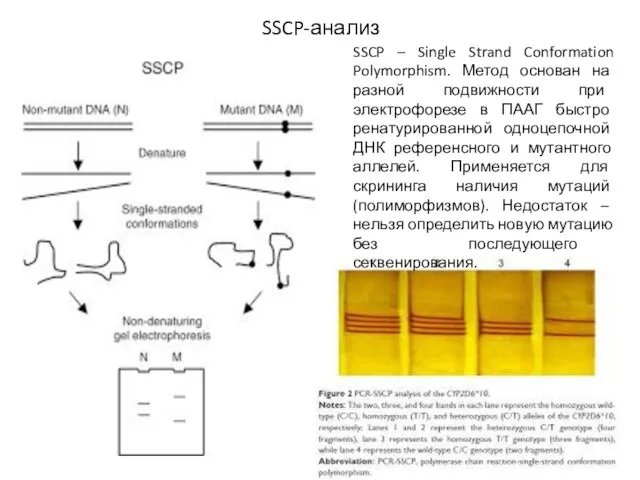
SSCP-анализ
SSCP – Single Strand Conformation Polymorphism. Метод основан на разной подвижности
SSCP-анализ
SSCP – Single Strand Conformation Polymorphism. Метод основан на разной подвижности

Мультиплексная ПЦР
При увеличении количества локусов в мультиплексной ПЦР, практически, экспоненциально растут
Мультиплексная ПЦР
При увеличении количества локусов в мультиплексной ПЦР, практически, экспоненциально растут
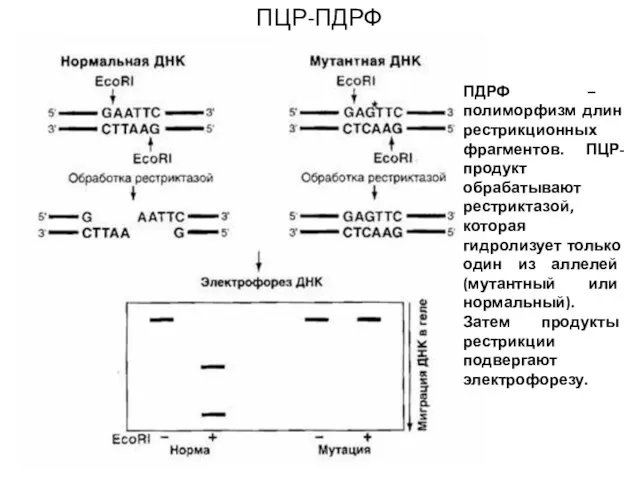
ПЦР-ПДРФ
ПДРФ – полиморфизм длин рестрикционных фрагментов. ПЦР-продукт обрабатывают рестриктазой, которая гидролизует
ПЦР-ПДРФ
ПДРФ – полиморфизм длин рестрикционных фрагментов. ПЦР-продукт обрабатывают рестриктазой, которая гидролизует
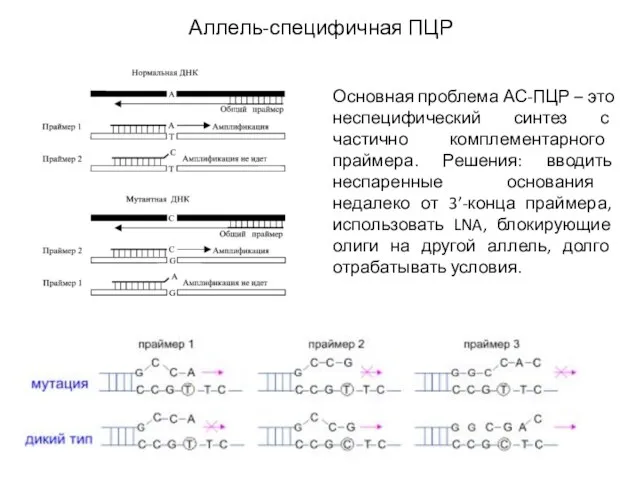
Аллель-специфичная ПЦР
Основная проблема АС-ПЦР – это неспецифический синтез с частично комплементарного
Аллель-специфичная ПЦР
Основная проблема АС-ПЦР – это неспецифический синтез с частично комплементарного
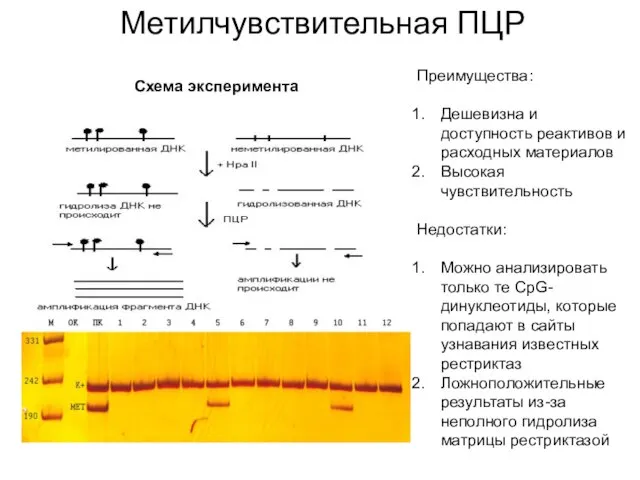
Метилчувствительная ПЦР
Схема эксперимента
Преимущества:
Дешевизна и доступность реактивов и расходных материалов
Высокая чувствительность
Недостатки:
Можно анализировать
Метилчувствительная ПЦР
Схема эксперимента
Преимущества:
Дешевизна и доступность реактивов и расходных материалов
Высокая чувствительность
Недостатки:
Можно анализировать
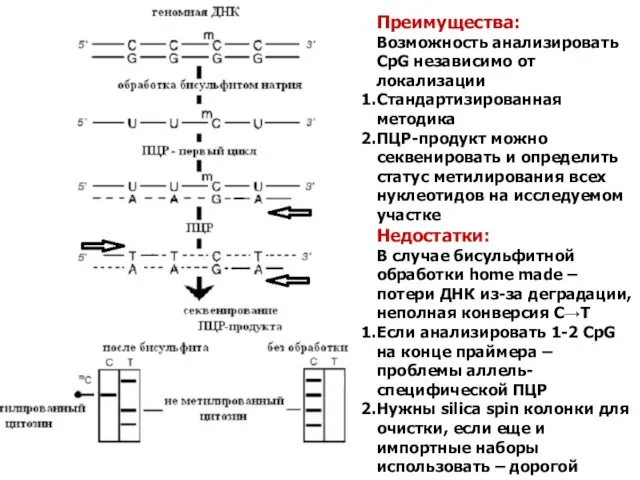
Преимущества:
Возможность анализировать CpG независимо от локализации
Стандартизированная методика
ПЦР-продукт можно секвенировать и определить
Преимущества:
Возможность анализировать CpG независимо от локализации
Стандартизированная методика
ПЦР-продукт можно секвенировать и определить
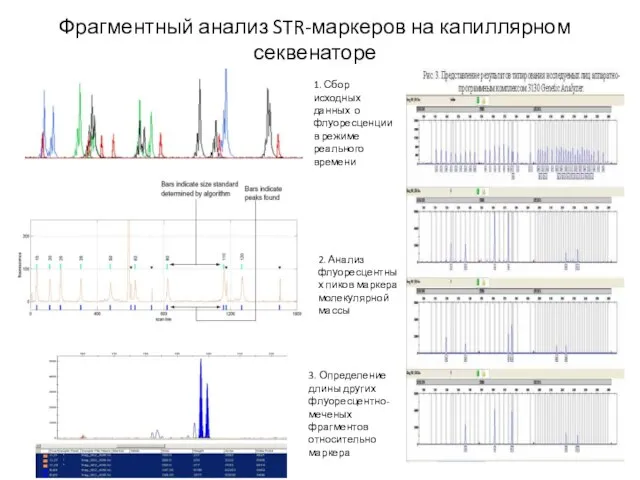
Фрагментный анализ STR-маркеров на капиллярном секвенаторе
1. Сбор исходных данных о флуоресценции
Фрагментный анализ STR-маркеров на капиллярном секвенаторе
1. Сбор исходных данных о флуоресценции
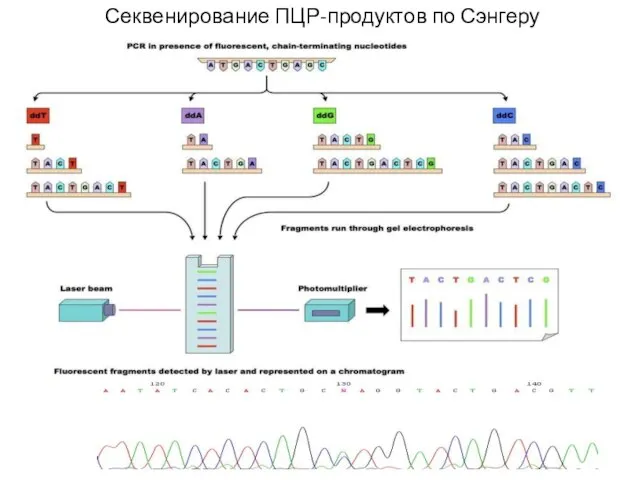
Секвенирование ПЦР-продуктов по Сэнгеру
Секвенирование ПЦР-продуктов по Сэнгеру
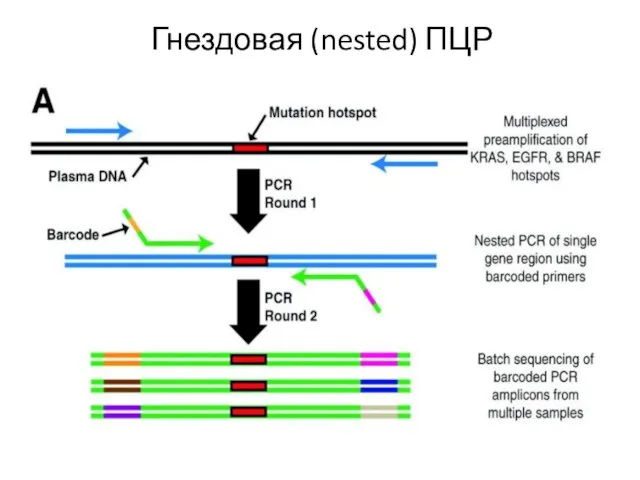
Гнездовая (nested) ПЦР
Гнездовая (nested) ПЦР
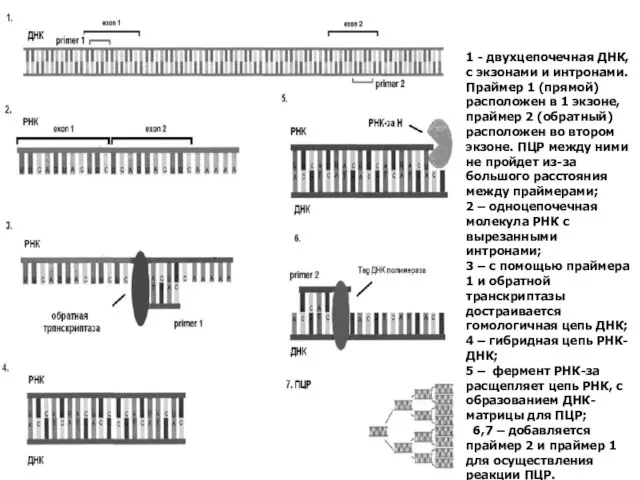
1 - двухцепочечная ДНК, с экзонами и интронами. Праймер 1 (прямой)
1 - двухцепочечная ДНК, с экзонами и интронами. Праймер 1 (прямой)

Hot-Start ПЦР
Hot-Start ПЦР
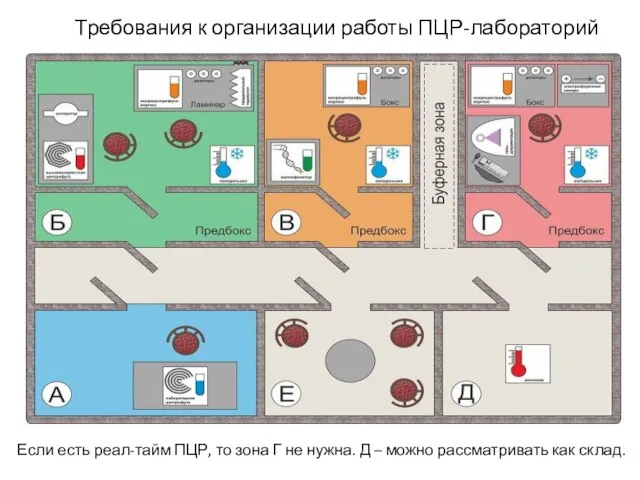
Требования к организации работы ПЦР-лабораторий
Если есть реал-тайм ПЦР, то зона Г
Требования к организации работы ПЦР-лабораторий
Если есть реал-тайм ПЦР, то зона Г
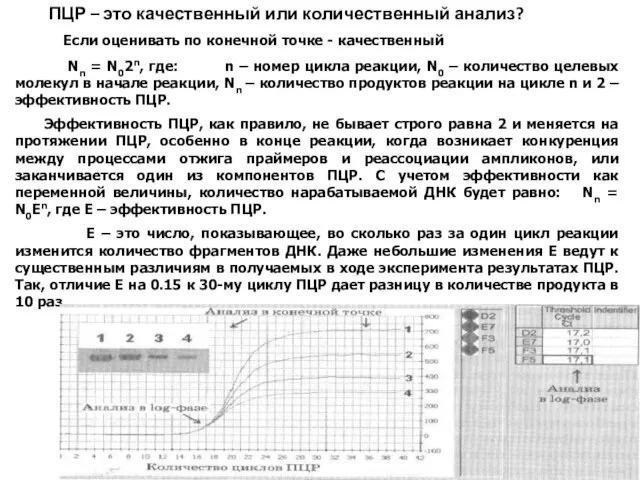
ПЦР – это качественный или количественный анализ?
Если оценивать по
ПЦР – это качественный или количественный анализ?
Если оценивать по

ПЦР – это качественный или количественный анализ? (продолжение)
Если измерять концентрации
ПЦР – это качественный или количественный анализ? (продолжение)
Если измерять концентрации
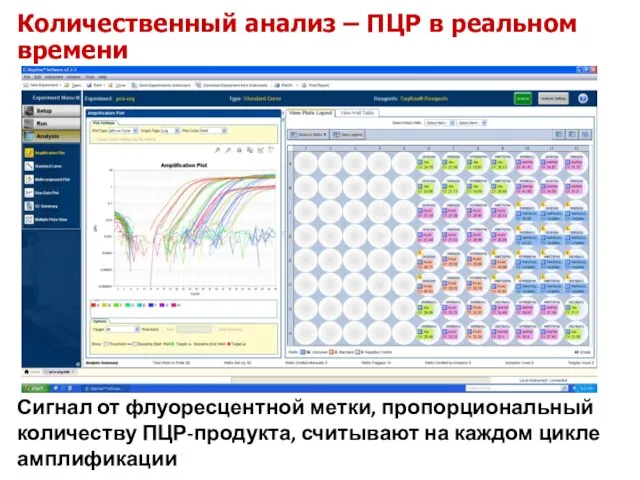
Количественный анализ – ПЦР в реальном времени
Сигнал от флуоресцентной метки, пропорциональный
Количественный анализ – ПЦР в реальном времени
Сигнал от флуоресцентной метки, пропорциональный
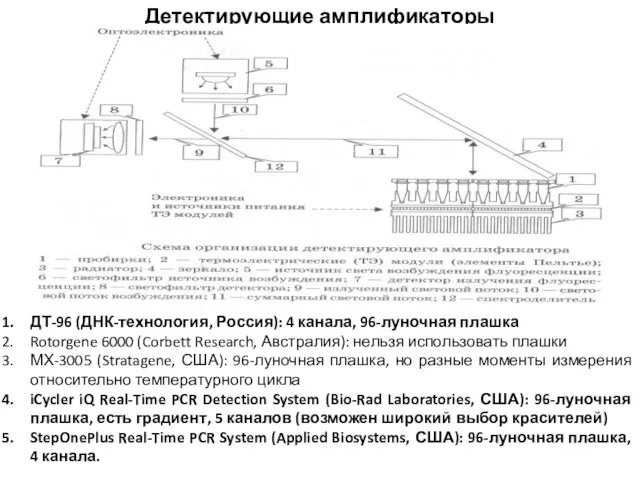
Детектирующие амплификаторы
ДТ-96 (ДНК-технология, Россия): 4 канала, 96-луночная плашка
Rotorgene 6000 (Corbett Research,
Детектирующие амплификаторы
ДТ-96 (ДНК-технология, Россия): 4 канала, 96-луночная плашка
Rotorgene 6000 (Corbett Research,

ПЦР в реальном времени (ПЦР-РВ)
Детектирующий амплификатор определяет количество ПЦР-продукта на каждом
ПЦР в реальном времени (ПЦР-РВ)
Детектирующий амплификатор определяет количество ПЦР-продукта на каждом

Пороговый метод сравнения графиков накопления ДНК
Суть порогового метода заключается в том,
Пороговый метод сравнения графиков накопления ДНК
Суть порогового метода заключается в том,
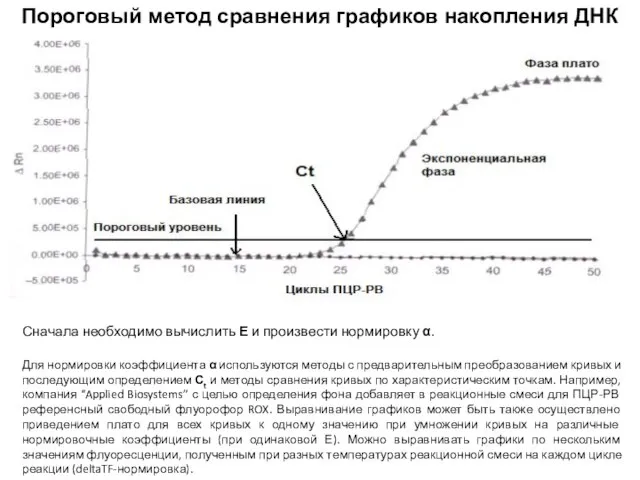
Пороговый метод сравнения графиков накопления ДНК
Сначала необходимо вычислить Е и произвести
Пороговый метод сравнения графиков накопления ДНК
Сначала необходимо вычислить Е и произвести
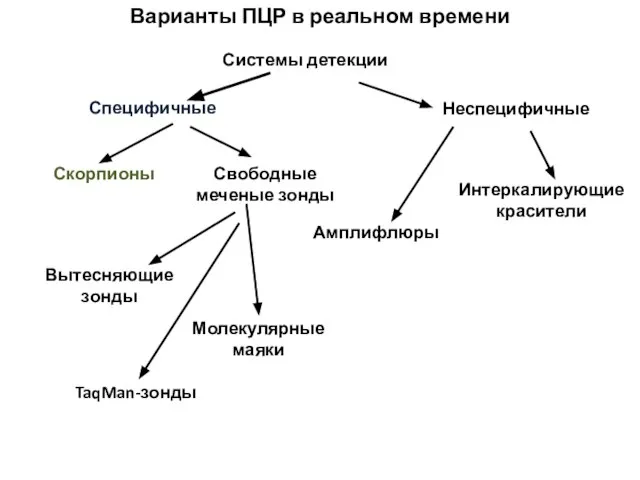
Варианты ПЦР в реальном времени
Системы детекции
Специфичные
Неспецифичные
Интеркалирующие красители
Амплифлюры
Скорпионы
Свободные меченые зонды
Вытесняющие зонды
TaqMan-зонды
Молекулярные маяки
Варианты ПЦР в реальном времени
Системы детекции
Специфичные
Неспецифичные
Интеркалирующие красители
Амплифлюры
Скорпионы
Свободные меченые зонды
Вытесняющие зонды
TaqMan-зонды
Молекулярные маяки
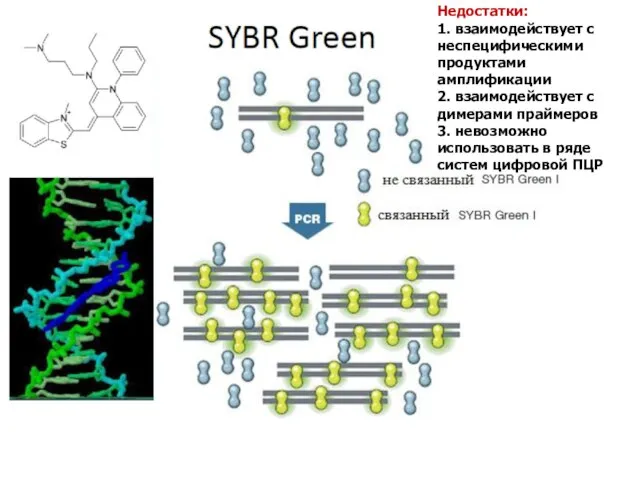
Недостатки:
1. взаимодействует с неспецифическими продуктами амплификации
2. взаимодействует с димерами праймеров
3. невозможно
Недостатки:
1. взаимодействует с неспецифическими продуктами амплификации
2. взаимодействует с димерами праймеров
3. невозможно

Амплифлюры
Амплифлюры
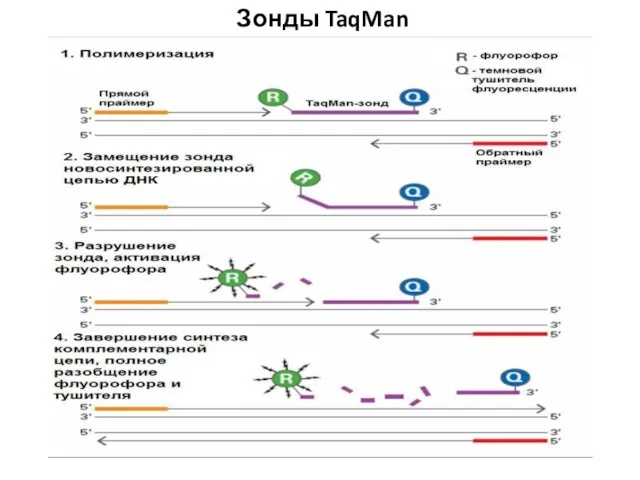
Зонды TaqMan
Зонды TaqMan

Молекулярный маяк представляет собой меченый красителем олигонуклеотид (25-40 нуклеотидов), который образует
Молекулярный маяк представляет собой меченый красителем олигонуклеотид (25-40 нуклеотидов), который образует
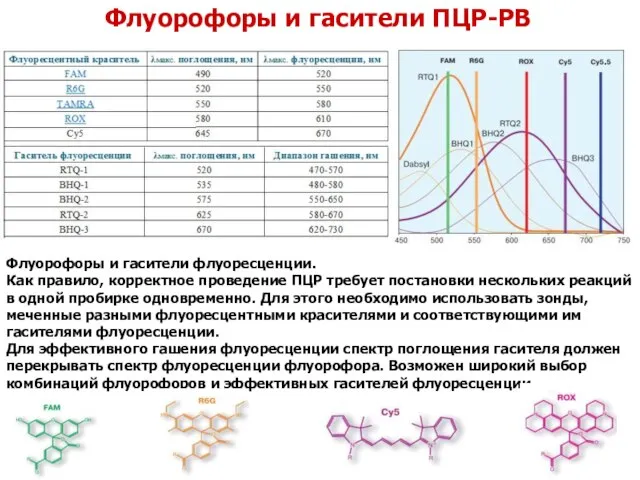
Флуорофоры и гасители ПЦР-РВ
Флуорофоры и гасители флуоресценции.
Как правило, корректное проведение ПЦР
Флуорофоры и гасители ПЦР-РВ
Флуорофоры и гасители флуоресценции.
Как правило, корректное проведение ПЦР
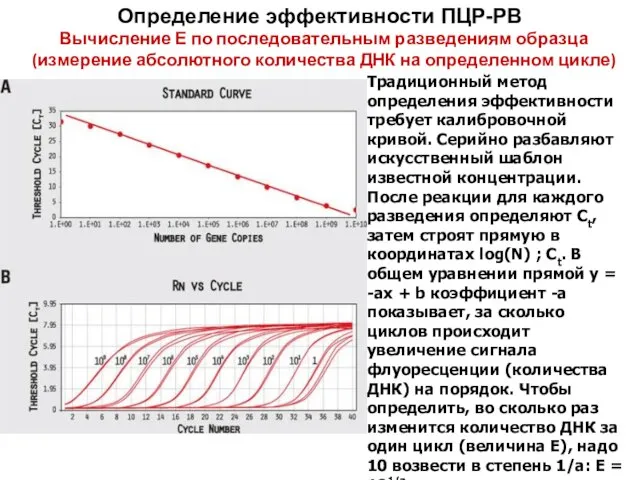
Определение эффективности ПЦР-РВ
Вычисление Е по последовательным разведениям образца
(измерение абсолютного количества ДНК
Определение эффективности ПЦР-РВ
Вычисление Е по последовательным разведениям образца
(измерение абсолютного количества ДНК
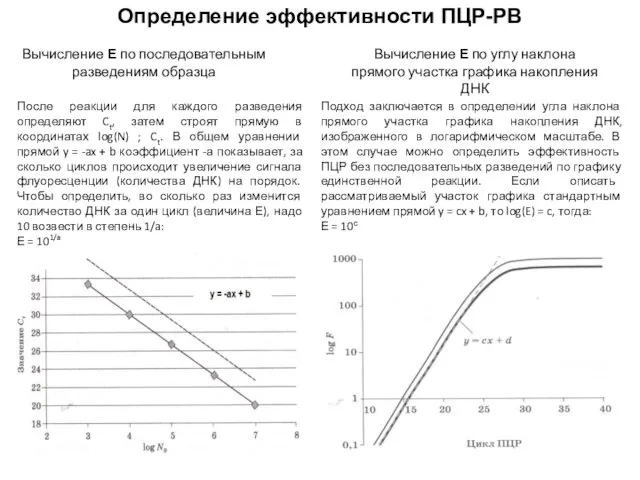
Определение эффективности ПЦР-РВ
Вычисление Е по последовательным разведениям образца
Вычисление Е по углу
Определение эффективности ПЦР-РВ
Вычисление Е по последовательным разведениям образца
Вычисление Е по углу

Абсолютное и относительное определение количества исходной ДНК
Формула N0 = N0rE(Ctr –
Абсолютное и относительное определение количества исходной ДНК
Формула N0 = N0rE(Ctr –

Абсолютное и относительное определение количества исходной ДНК (продолжение)
Предполагая, что условия ПЦР-РВ
Абсолютное и относительное определение количества исходной ДНК (продолжение)
Предполагая, что условия ПЦР-РВ
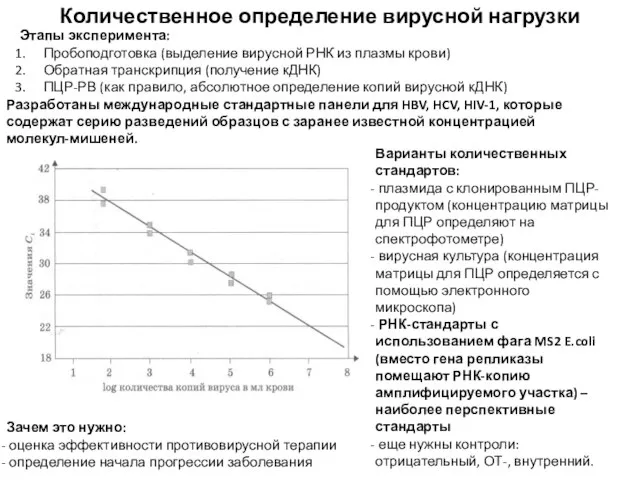
Количественное определение вирусной нагрузки
Этапы эксперимента:
Пробоподготовка (выделение вирусной РНК из плазмы крови)
Обратная
Количественное определение вирусной нагрузки
Этапы эксперимента:
Пробоподготовка (выделение вирусной РНК из плазмы крови)
Обратная

ДНК-диагностика онкогематологических заболеваний
Полная ремиссия: отсутствие клинических симптомов болезни (клиническая ремиссия) и
ДНК-диагностика онкогематологических заболеваний
Полная ремиссия: отсутствие клинических симптомов болезни (клиническая ремиссия) и
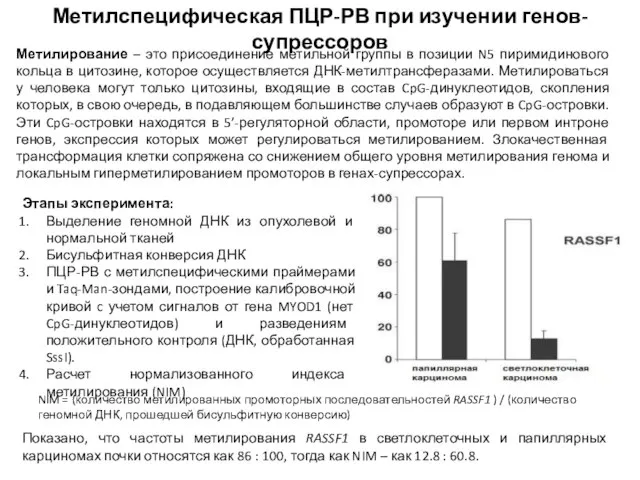
Метилспецифическая ПЦР-РВ при изучении генов-супрессоров
Метилирование – это присоединение метильной группы в
Метилспецифическая ПЦР-РВ при изучении генов-супрессоров
Метилирование – это присоединение метильной группы в
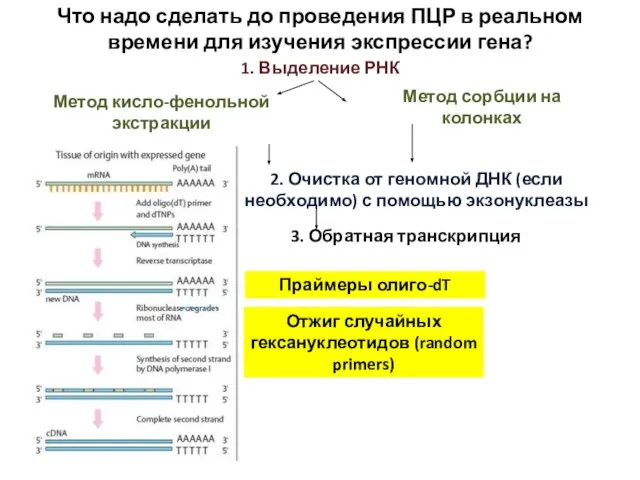
Что надо сделать до проведения ПЦР в реальном времени для изучения
Что надо сделать до проведения ПЦР в реальном времени для изучения
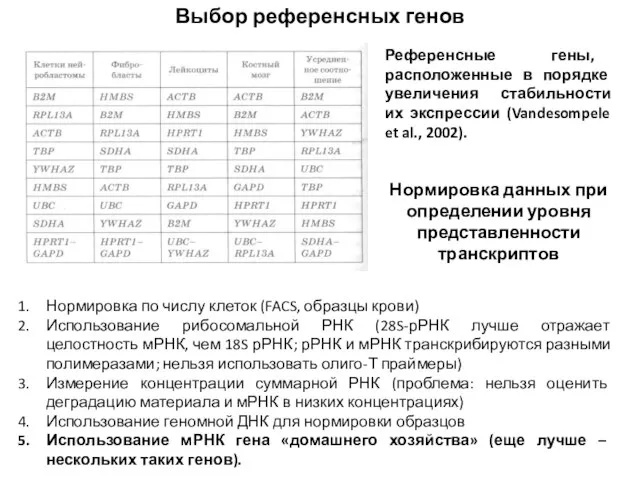
Выбор референсных генов
Референсные гены, расположенные в порядке увеличения стабильности их экспрессии
Выбор референсных генов
Референсные гены, расположенные в порядке увеличения стабильности их экспрессии
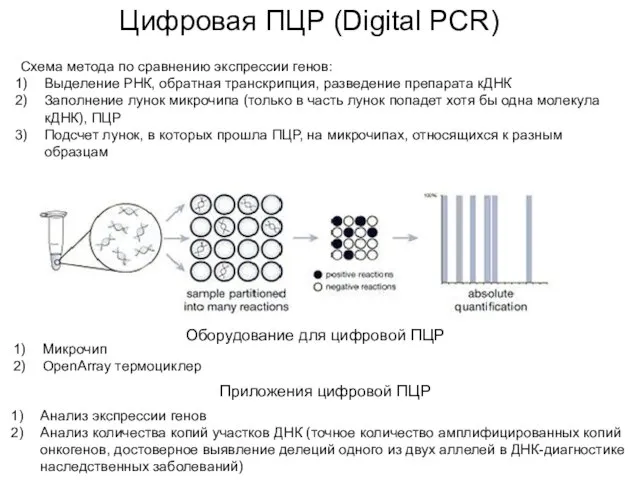
Цифровая ПЦР (Digital PCR)
Схема метода по сравнению экспрессии генов:
Выделение РНК, обратная
Цифровая ПЦР (Digital PCR)
Схема метода по сравнению экспрессии генов:
Выделение РНК, обратная

Выходящий тестовый контроль
1. Увеличение количества ПЦР-продуктов при эффективности ПЦР, равной 2,
Выходящий тестовый контроль
1. Увеличение количества ПЦР-продуктов при эффективности ПЦР, равной 2,

 Глобальная проблема коронавируса и его последствия
Глобальная проблема коронавируса и его последствия Перитонит. Клиническая картина и диагностика
Перитонит. Клиническая картина и диагностика Введение в клиническую биохимию. Биологические маркеры
Введение в клиническую биохимию. Биологические маркеры Покровные протезы
Покровные протезы Trypanosoma Brucei
Trypanosoma Brucei Общество православных студентов-медиков СГМУ
Общество православных студентов-медиков СГМУ АО ННМЦ Кетамин
АО ННМЦ Кетамин Токсическое действие алкоголя на организм человека
Токсическое действие алкоголя на организм человека Основы реабилитации пациентов. Тема 1
Основы реабилитации пациентов. Тема 1 SARS-CoV-2. Тяжёлый острый респираторный синдром – коронавирус 2 COVID – 19
SARS-CoV-2. Тяжёлый острый респираторный синдром – коронавирус 2 COVID – 19 Врожденная непроходимость желудочно-кишечного тракта
Врожденная непроходимость желудочно-кишечного тракта Гистология
Гистология Атеросклероз. Этиологисы
Атеросклероз. Этиологисы Парапроктит и свищи прямой кишки
Парапроктит и свищи прямой кишки Дифференциальная диагностика: патопсихологические симптомокомплексы
Дифференциальная диагностика: патопсихологические симптомокомплексы Аллергия. Типы гиперчувствительности
Аллергия. Типы гиперчувствительности Штифт или вкладка: что лучше выбрать под зубную коронку
Штифт или вкладка: что лучше выбрать под зубную коронку Черепно-мозговые нервы III - V. Методика исследования. Симптомы поражения. Альтернирующие синдромы. (Занятие 4)
Черепно-мозговые нервы III - V. Методика исследования. Симптомы поражения. Альтернирующие синдромы. (Занятие 4) Жедел пневмониялар: бронхопневмония, крупозды, аралық. Морфологиялық сипаттамасы, нәтижелері және асқынулары
Жедел пневмониялар: бронхопневмония, крупозды, аралық. Морфологиялық сипаттамасы, нәтижелері және асқынулары Мімічна мускулатура
Мімічна мускулатура Спинномозговые нервы
Спинномозговые нервы Основы физиотерапии. Тема 3
Основы физиотерапии. Тема 3 Ембріотехнології. Виникнення ембріотехнологій
Ембріотехнології. Виникнення ембріотехнологій Ритмокардиография в спорте
Ритмокардиография в спорте Антиредукционизм в отечественной психологии и в работах В.П. Зинченко
Антиредукционизм в отечественной психологии и в работах В.П. Зинченко Воспаления среднего уха у новорожденных и грудных детей
Воспаления среднего уха у новорожденных и грудных детей Точечный массаж
Точечный массаж Использование углерода в медицине
Использование углерода в медицине