- Онкогенетика

Содержание
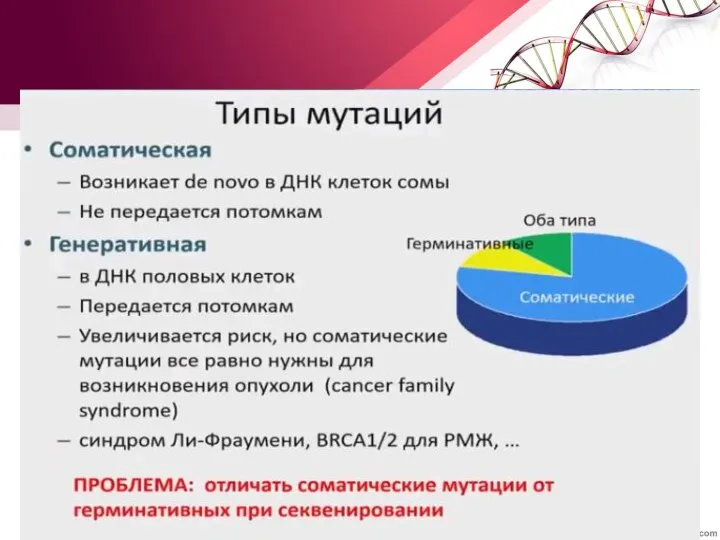
- 2. ОНКОГЕНЕТИКА направление генетики, изучающее причины и законы возникновения и функционирования опухоли.
- 3. ОПУХОЛИ Доброкачественные: Экспансивный медленный рост; Не прорастают в окружающие ткани; Имеют капсулу, отграничивающую опухоль от окружающих
- 4. ОПУХОЛЬ Новообразование, неоплазма - патологическое образование, характеризующееся неконтролируемым размножением клеток с нарушением их роста и дифференцировки,
- 5. Опухоли Саркомы (опухоль возникает в мезенхимальной ткани, например в костях, мышцах, соединительной ткани или в тканях
- 6. Основные причины смерти в популяции (по данным ВОЗ)
- 7. КАНЦЕРОГЕНЫ физические ультрафиолетовое и ионизирующее излучение; химические асбест, компоненты табачного дыма, афлатоксины (загрязнители пищевых продуктов) мышьяк
- 8. КАНЦЕРОГЕНЕЗ Канцерогенез — сложный патофизиологический процесс зарождения и развития опухоли. (син. онкогенез). Канцерогенез — сложный многоэтапный
- 9. Генетические аспекты канцерогенеза Генетический аппарат клеток обладает сложной системой контроля деления, роста и дифференцировки клеток Протоонкогены
- 10. Гены-супрессоры опухолей Функции генов-супрессоров противоположны функциям протоонкогенов. Гены-супрессоры оказывают тормозящее влияние на процессы клеточного деления и
- 11. Онкогены – мутировавшие протоонкогены Протоонкогены – в нормальных клетках стимулируют клеточное деление Примеры (EGFR, KRAS, MET,
- 12. .
- 13. Опухолевые супрессоры Gatekeepers (хранители клеточного цикла) - Контролируют клеточный рост и деление, останавливая клеточный цикл, вызывают
- 14. Клеточный цикл G1 – пресинтетический период S – синтетический период Автокаталитическая G2 – постсинтетический период интерфаза
- 15. Митотическая активность клеток различных тканей Постоянно делящиеся в обновляющихся тканях (клетки базального слоя покровного эпителия, в
- 16. АПОПТОЗ генетически запрограммированная гибель клеток многоклеточного организма. Роль апоптоза: необходим для нормального формирования органов в онтогенезе,
- 17. В норме апоптоз начинается с активации так называемых «рецепторов смерти» на поверхности мембраны клетки путем присоединения
- 18. Индукция апоптоза
- 19. Внешний путь активации апоптоза Внешний путь начинается со связывания CD95 со своим лигандом, CD95L. В результате
- 20. Внутренний путь активации апоптоза Внутренний путь апоптоза запускается различными воздействиями (отсутствие ростовых факторов, стресс и повреждение).
- 22. Современная двухударная модель канцерогенеза
- 23. Хромотрипсис
- 24. Хромоплексия
- 25. Сравнение хромотрипсиса и хромоплексии
- 26. Категис
- 27. Эволюционная теория Ч.Дарвина Раскрывает пути и закономерности эволюционного процесса видообразования Впоследствии она оказалась применима для других
- 28. Гетерогенность опухолевых клеток Существуют две основные гипотезы происхождения гетерогенности опухолевых клеток: различные субклоны опухолевых клеток возникают
- 29. Гетерогенность опухолевых клеток Естественная клональная эволюция происходит за счет внутренних механизмов и определяется драйверными мутациями (возникающими
- 30. Прогрессирование рака
- 31. Прогрессирование рака Constant population
- 32. Прогрессирование рака Advantageous mutant
- 33. Clonal expansion Прогрессирование рака
- 34. Saturation Прогрессирование рака
- 35. Advantageous mutant Прогрессирование рака
- 36. Wave of clonal expansion Прогрессирование рака
- 38. Борьба с гетерогенностью - Найти (targetable) события, которые произошли на ранней стадии и поэтому есть у
- 39. Мутации драйверы и мутации пассажиры
- 40. Возникающие генетические изменения в опухолевых клетках не всегда приводят к функциональным последствиям, часть из них нейтральны.
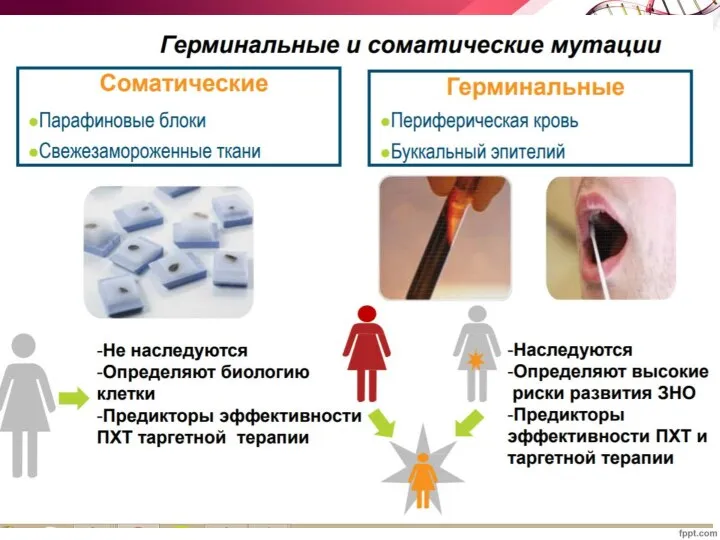
- 43. Генетические нарушения Делеции целых хромосомных районов, содержащих гены супрессоры опухолевого роста (потеря гетерозиготности); Дупликации или амплификации
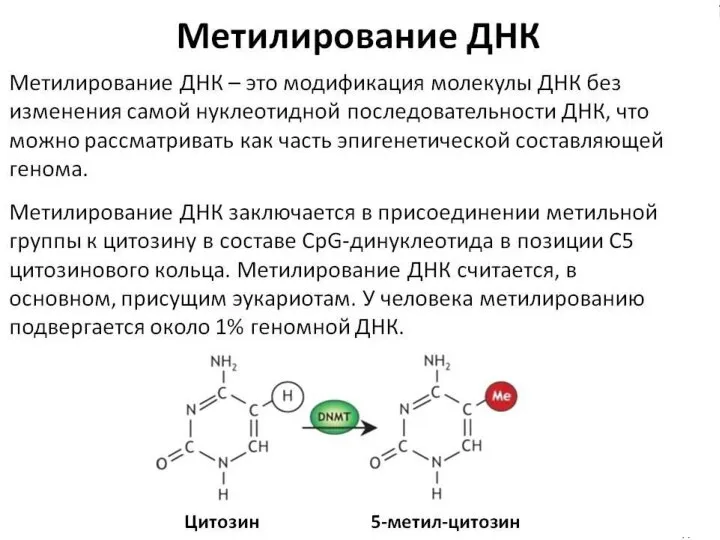
- 44. ЭПИГЕНЕТИЧЕСКИЕ ФАКТОРЫ наследственные и ненаследственные изменения в экспрессии конкретного гена без каких-либо соответствующих структурных изменений в
- 46. Аномально метилированные промоторные районы (CpG-островки) составляют 100-400 в каждой опухоли.
- 47. Метилирование ДНК является ценным биомаркером для диагностики рака 1. Целый ряд генов, прямо или косвенно вовлеченных
- 48. Частоты метилирования генов-супрессоров при раке почки В опухолях метилирование VHL определено в 14.2%, RASSF1 – 52.8%,
- 49. Методы анализа метилирования 1. Метилчувствительная ПЦР (NotI, EagI, SacII, HpaII, HhaI) аналитическая чувствительность - 1: 2000
- 50. Системы молекулярных маркеров метилирования
- 51. Метилирование промоторных районов генов RASSF и р16 достоверно чаще происходит в клетках уротелиальных карцином с инвазией
- 52. Метилирование позволяет предсказать поведение опухоли (эффективность терапии, метастазирование): - метилирование WIT1 коррелирует с хеморезистентностью при ОМЛ;
- 53. Метилирование ДНК, как диагностический маркер онкологического заболевания, имеет ряд преимуществ перед другими маркерами Мы определяем положительный
- 54. Молекулярные маркеры, определяемые на ранних стадиях канцерогенеза.
- 55. Метилирование позволяет более точно классифицировать типы и подтипы опухолей: - гиперметилирование BRCA1 обнаружено при раке молочной
- 56. Структура хроматина и модификация гистонов (James Larkin et al., 2012)
- 57. Карта метилирования и ацетилирования гистонов при почечно-клеточных карциномах (James Larkin et al., 2012)
- 58. РНК-интерференция: процесс подавления экспрессии гена на стадии транскрипции, трансляции, деаденилирования ил деградации мРНК при помощи малых
- 59. МикроРНК - некодирующие РНК, состоящие из 18-24 нуклеотидов и регулирующие экспрессиюдо 30% генов человека. Микро РНК
- 60. Чиповые технологии для высокопроизводительного генотипирования Affymetrix Agilent technologies Genepix Asper Biotech Perkin Elmer Illumina
- 61. Белок р53 Фактор апоптоза p53 определяет повреждения ДНК останавливает синтез ДНК до тех пор, пока не
- 63. Белок р53 Синдром Ли Фраумени Наследственная предрасположенность к раку с вероятностью 90% до возраста 60 лет
- 64. Ретинобластома Злокачественная опухоль глаза, развивается преимущественно в детском возрасте из тканей эмбрионального происхождения. Диагностируется в возрасте
- 65. Ретинобластома Наследственная: Аутосомно-доминантный тип наследования У носителей гена RB 90% шанс развития ретинобластомы Опухоли в обоих
- 66. Ретинобластома Спорадическая: Мутации в обеих копиях гена RB1 Опухоль поражает только один глаз Риск опухолей другой
- 67. Ген RB1 Расположен на хромосоме 13q14 и кодирует белок pRB pRB – негативный регулятор клеточного цикла
- 68. Рак молочной железы В Европе ежегодно диагностируется 430000 новых случаев рака груди ~5% у лиц с
- 69. Критерии для постановки диагноза наследственного рака молодой возраст возникновения онкологического заболевания; Наследственный анамнез (наличие в семье
- 70. BRCA1 Повышенный риск других видов рака Рак груди 56%-87% (часто с ранним началом) Второй первичный рак
- 71. BRCA2 Другие виды рака: поджелудочной железы, меланома Рак груди (50%-80%) Рак яичников (15-27%) Рак груди (7%)
- 72. Экспрессия повышена в точке перехода из фазы G1 в фазу S и в течение фазы S
- 73. Скрининговые обследования: Маммография/МРТ Профилактическая мастэктомия ~90% снижение риска Профилактическая двусторонняя сальпингооофория ~96-98% снижение риска рака яичников
- 75. Показания: 2 родственников первой или второй степени с раком груди в возрасте до 50 лет 3
- 76. BRCA1 – ассоциированные опухоли – подходы к терапии - Пациенты хорошо отвечают на лечение фармакологическими ингибиторами
- 77. Тактика ведения пациентов с
- 79. Наследственный рак предстательной железы
- 80. Ключевые молекулярные маркеры рака предстательной железы Эпигенетические изменения Метилирование генов p16, HIC1, N33 и GSTP1 в
- 84. Скачать презентацию

ОНКОГЕНЕТИКА
направление генетики, изучающее причины и законы возникновения и функционирования опухоли.
ОНКОГЕНЕТИКА
направление генетики, изучающее причины и законы возникновения и функционирования опухоли.

ОПУХОЛИ
Доброкачественные:
Экспансивный медленный рост;
Не прорастают в окружающие ткани;
Имеют капсулу, отграничивающую опухоль от
ОПУХОЛИ
Доброкачественные:
Экспансивный медленный рост;
Не прорастают в окружающие ткани;
Имеют капсулу, отграничивающую опухоль от

ОПУХОЛЬ
Новообразование, неоплазма - патологическое образование, характеризующееся неконтролируемым размножением клеток с нарушением
ОПУХОЛЬ
Новообразование, неоплазма - патологическое образование, характеризующееся неконтролируемым размножением клеток с нарушением

Опухоли
Саркомы (опухоль возникает в мезенхимальной ткани, например в костях, мышцах, соединительной
Опухоли
Саркомы (опухоль возникает в мезенхимальной ткани, например в костях, мышцах, соединительной
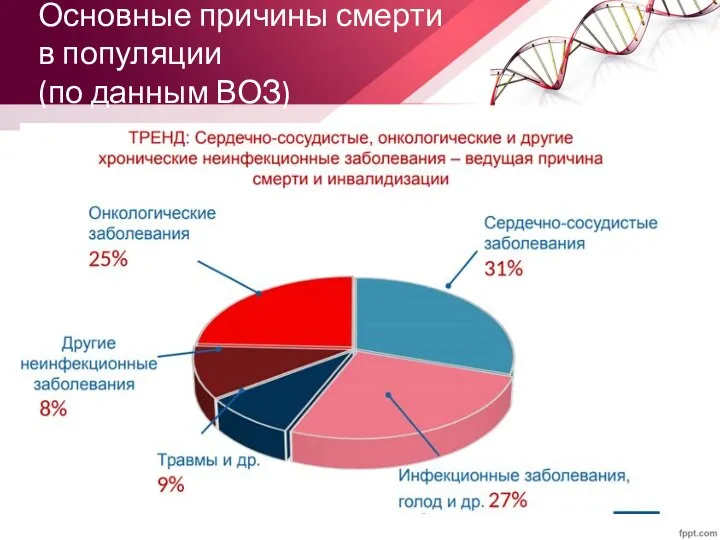
Основные причины смерти
в популяции
(по данным ВОЗ)
Основные причины смерти
в популяции
(по данным ВОЗ)

КАНЦЕРОГЕНЫ
физические
ультрафиолетовое и ионизирующее излучение;
химические
асбест,
компоненты табачного дыма,
афлатоксины (загрязнители
КАНЦЕРОГЕНЫ
физические
ультрафиолетовое и ионизирующее излучение;
химические
асбест,
компоненты табачного дыма,
афлатоксины (загрязнители

КАНЦЕРОГЕНЕЗ
Канцерогенез — сложный патофизиологический процесс зарождения и развития опухоли. (син. онкогенез).
Канцерогенез — сложный многоэтапный
КАНЦЕРОГЕНЕЗ
Канцерогенез — сложный патофизиологический процесс зарождения и развития опухоли. (син. онкогенез).
Канцерогенез — сложный многоэтапный

Генетические аспекты канцерогенеза
Генетический аппарат клеток обладает сложной системой контроля деления, роста
Генетические аспекты канцерогенеза
Генетический аппарат клеток обладает сложной системой контроля деления, роста

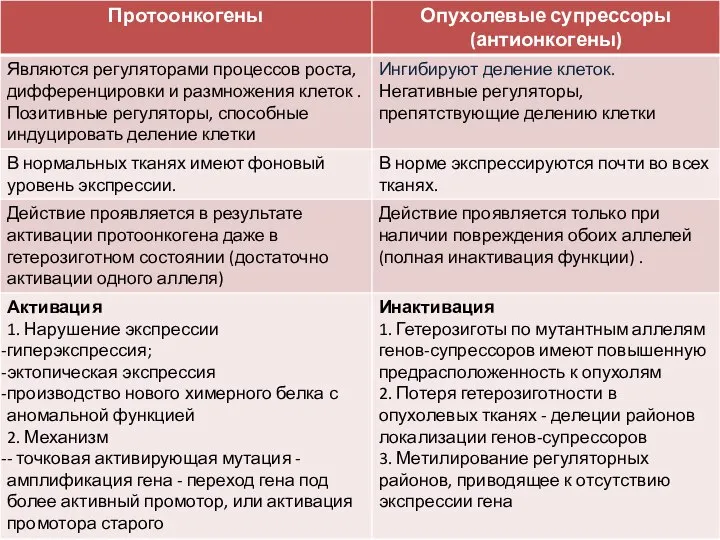
Гены-супрессоры опухолей
Функции генов-супрессоров противоположны функциям протоонкогенов. Гены-супрессоры оказывают тормозящее влияние на процессы клеточного деления и
Гены-супрессоры опухолей
Функции генов-супрессоров противоположны функциям протоонкогенов. Гены-супрессоры оказывают тормозящее влияние на процессы клеточного деления и
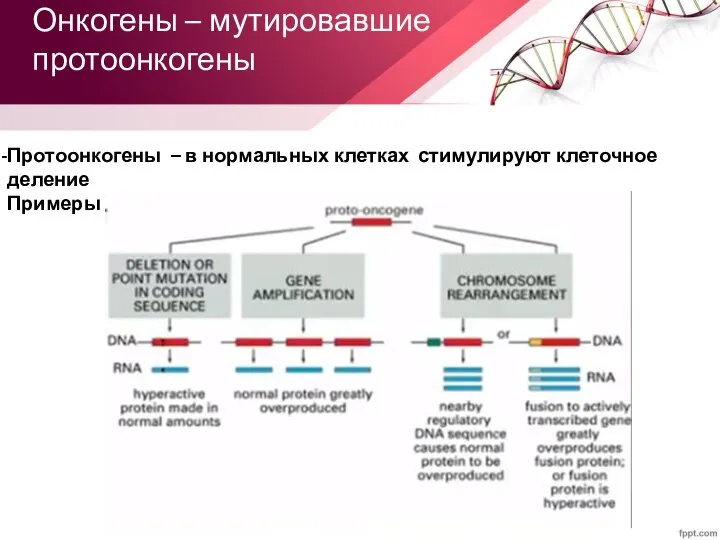
Онкогены – мутировавшие
протоонкогены
Протоонкогены – в нормальных клетках стимулируют клеточное деление
Примеры
Онкогены – мутировавшие
протоонкогены
Протоонкогены – в нормальных клетках стимулируют клеточное деление
Примеры
.
.
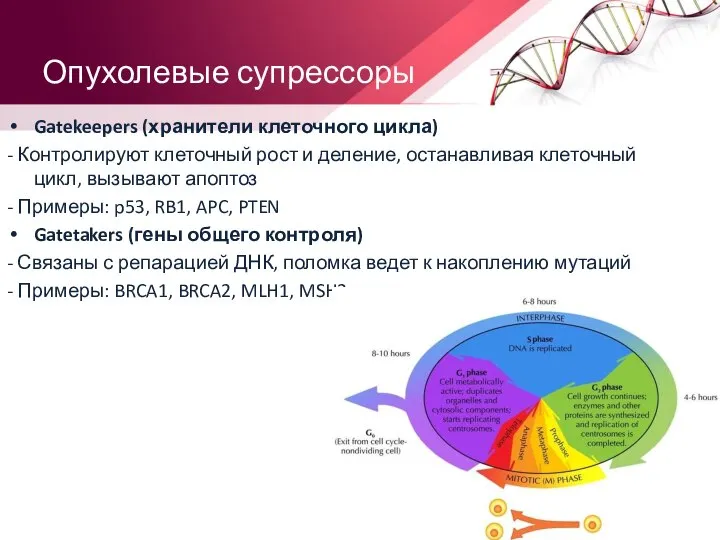
Опухолевые супрессоры
Gatekeepers (хранители клеточного цикла)
- Контролируют клеточный рост и деление, останавливая
Опухолевые супрессоры
Gatekeepers (хранители клеточного цикла)
- Контролируют клеточный рост и деление, останавливая
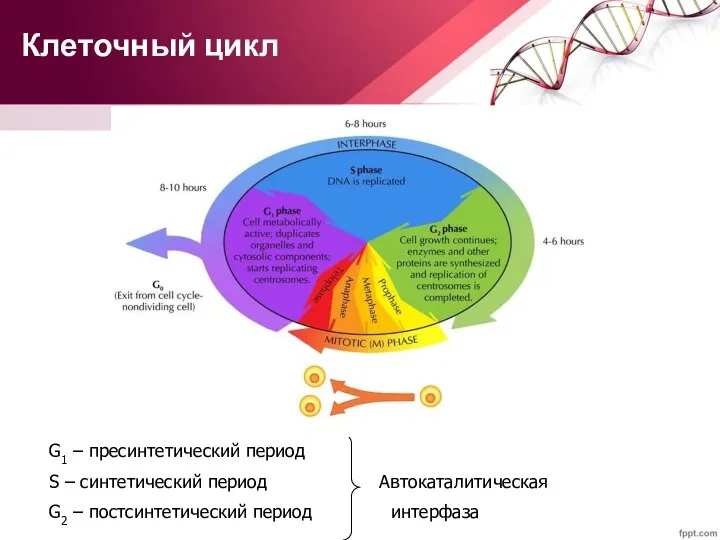
Клеточный цикл
G1 – пресинтетический период
S – синтетический период Автокаталитическая
G2 – постсинтетический
Клеточный цикл
G1 – пресинтетический период
S – синтетический период Автокаталитическая
G2 – постсинтетический

Митотическая активность клеток различных тканей
Постоянно делящиеся в обновляющихся тканях (клетки базального
Митотическая активность клеток различных тканей
Постоянно делящиеся в обновляющихся тканях (клетки базального

АПОПТОЗ
генетически запрограммированная гибель клеток многоклеточного организма.
Роль апоптоза:
необходим для нормального формирования
АПОПТОЗ
генетически запрограммированная гибель клеток многоклеточного организма.
Роль апоптоза:
необходим для нормального формирования

В норме апоптоз начинается с активации так называемых «рецепторов смерти» на
В норме апоптоз начинается с активации так называемых «рецепторов смерти» на
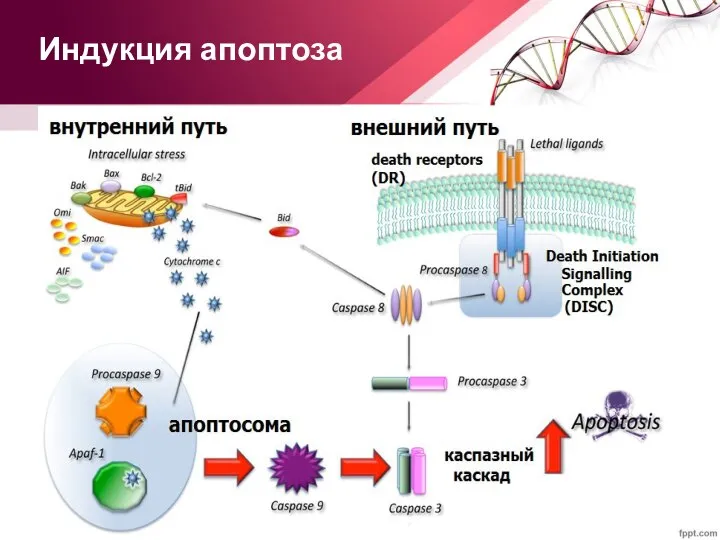
Индукция апоптоза
Индукция апоптоза

Внешний путь активации апоптоза
Внешний путь начинается со связывания CD95 со
Внешний путь активации апоптоза
Внешний путь начинается со связывания CD95 со

Внутренний путь активации апоптоза
Внутренний путь апоптоза запускается различными воздействиями (отсутствие
Внутренний путь активации апоптоза
Внутренний путь апоптоза запускается различными воздействиями (отсутствие
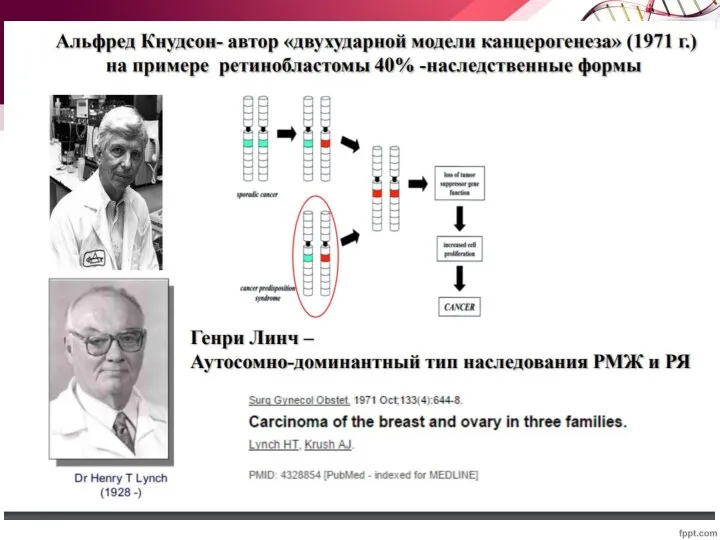
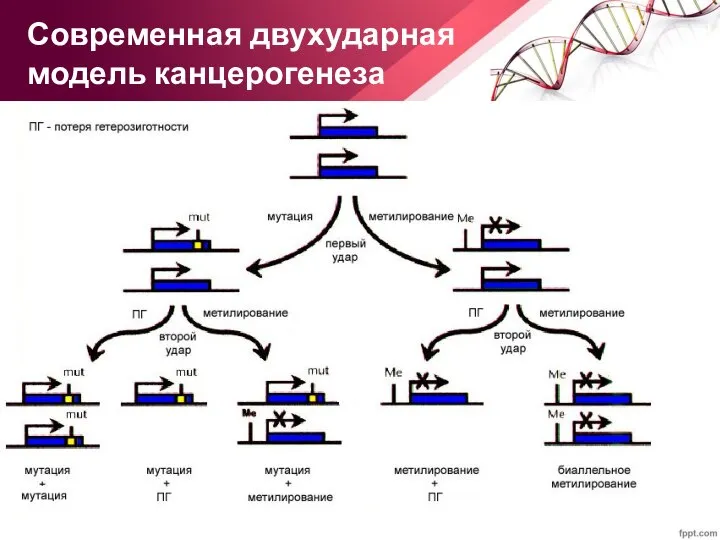
Современная двухударная модель канцерогенеза
Современная двухударная модель канцерогенеза
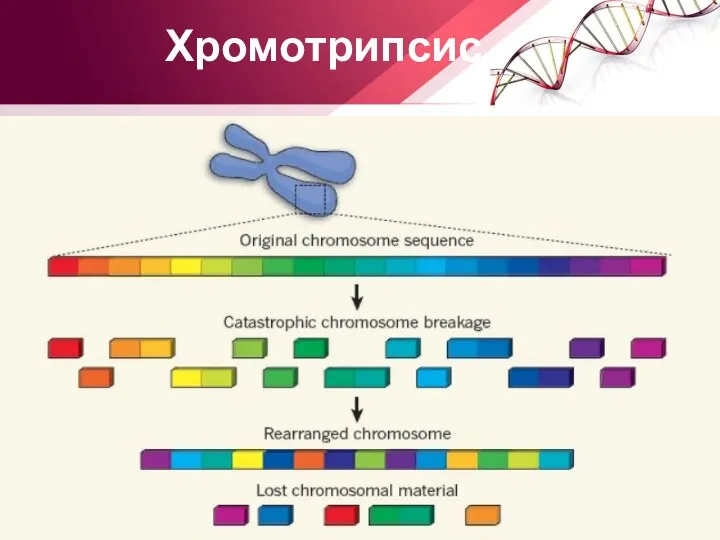
Хромотрипсис
Хромотрипсис
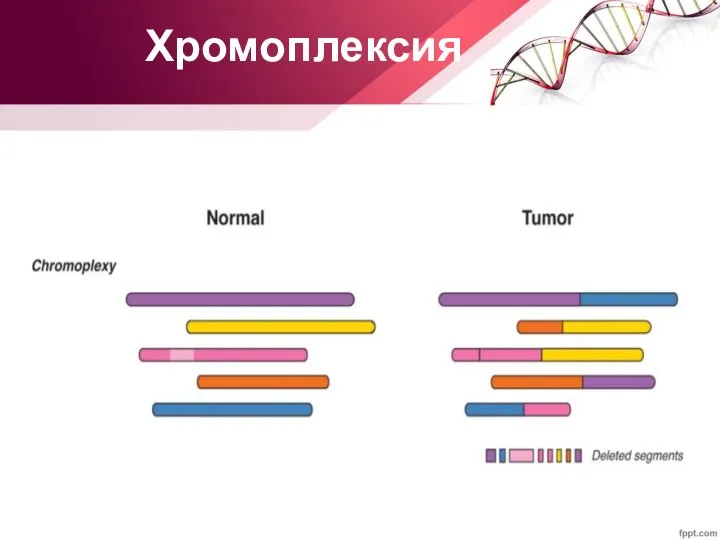
Хромоплексия
Хромоплексия
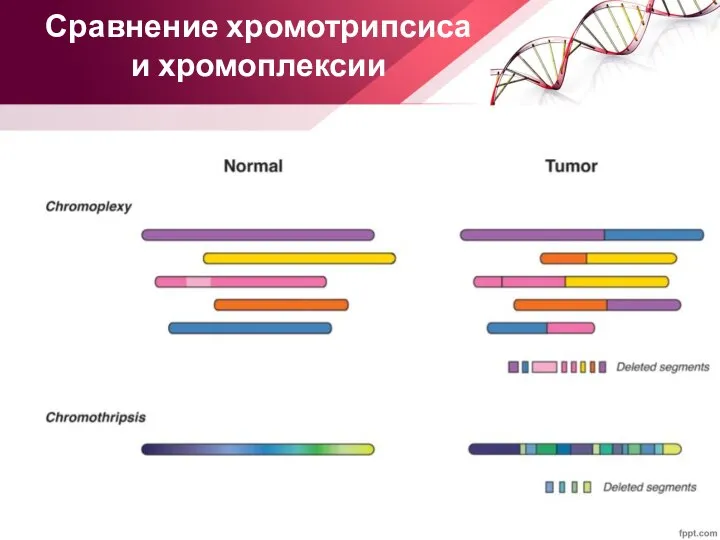
Сравнение хромотрипсиса
и хромоплексии
Сравнение хромотрипсиса
и хромоплексии
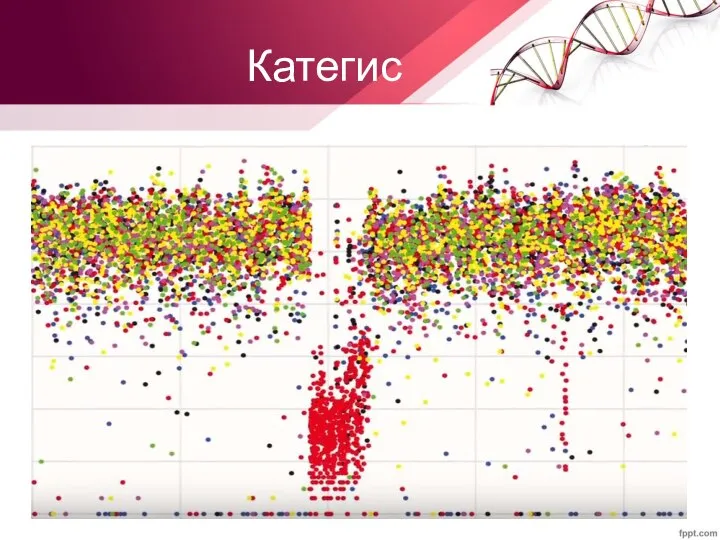
Категис
Категис
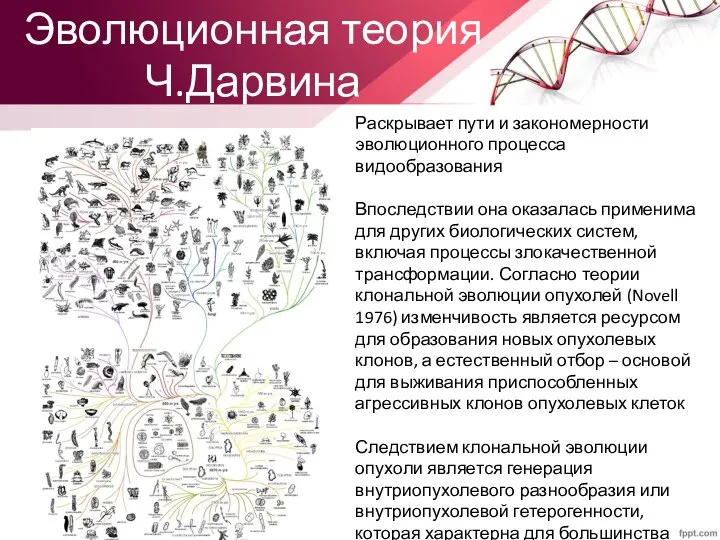
Эволюционная теория
Ч.Дарвина
Раскрывает пути и закономерности эволюционного процесса видообразования
Впоследствии она оказалась
Эволюционная теория
Ч.Дарвина
Раскрывает пути и закономерности эволюционного процесса видообразования
Впоследствии она оказалась

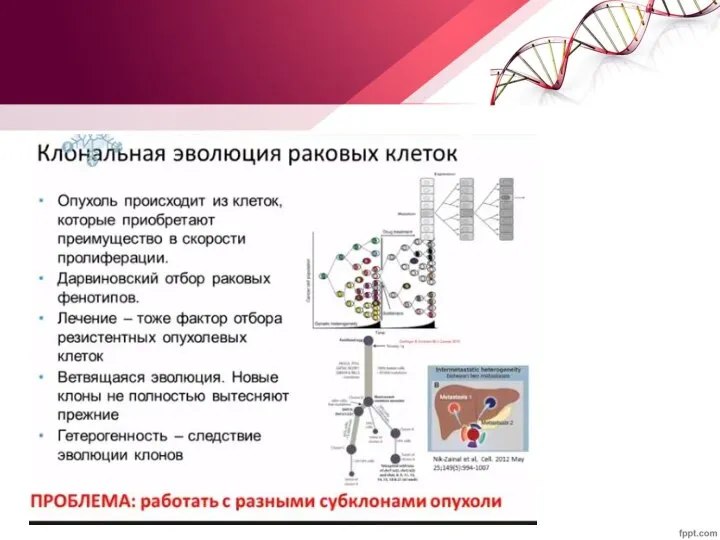
Гетерогенность опухолевых клеток
Существуют две основные гипотезы происхождения гетерогенности опухолевых клеток:
различные
Гетерогенность опухолевых клеток
Существуют две основные гипотезы происхождения гетерогенности опухолевых клеток:
различные

Гетерогенность опухолевых клеток
Естественная клональная эволюция происходит за счет внутренних механизмов и
Гетерогенность опухолевых клеток
Естественная клональная эволюция происходит за счет внутренних механизмов и

Прогрессирование
рака
Прогрессирование
рака

Прогрессирование
рака
Constant population
Прогрессирование
рака
Constant population
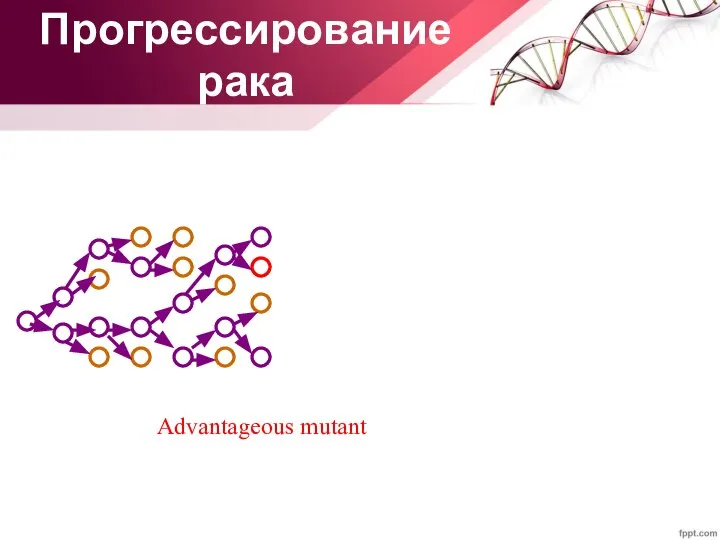
Прогрессирование
рака
Advantageous mutant
Прогрессирование
рака
Advantageous mutant
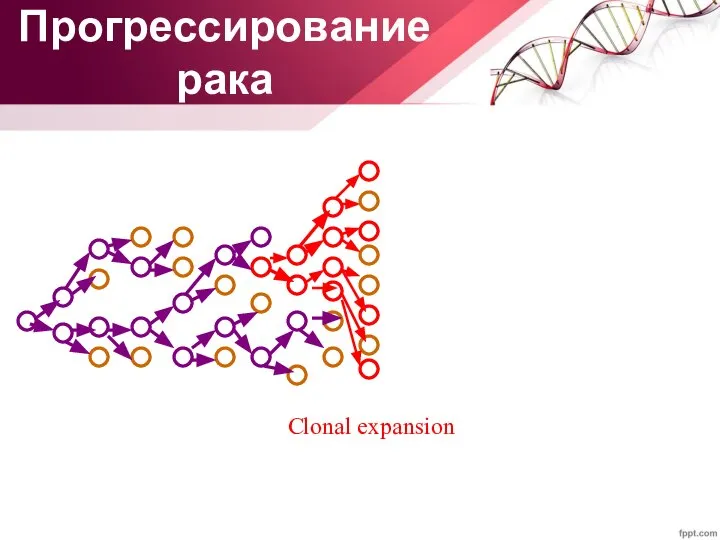
Clonal expansion
Прогрессирование
рака
Clonal expansion
Прогрессирование
рака
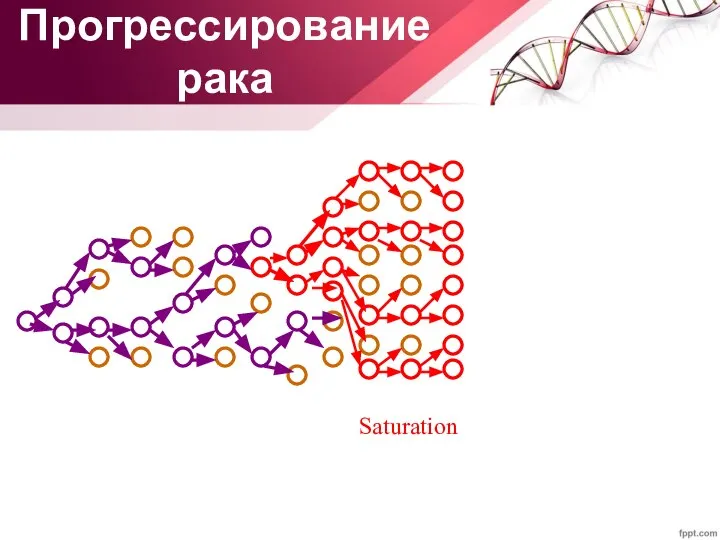
Saturation
Прогрессирование
рака
Saturation
Прогрессирование
рака
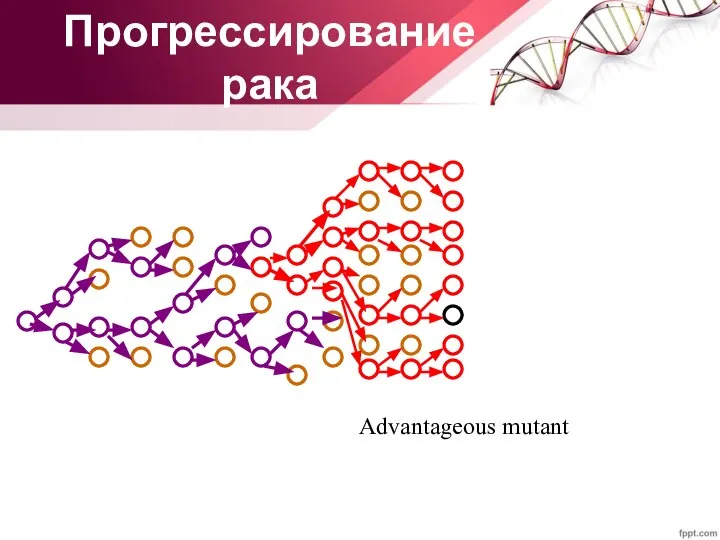
Advantageous mutant
Прогрессирование
рака
Advantageous mutant
Прогрессирование
рака
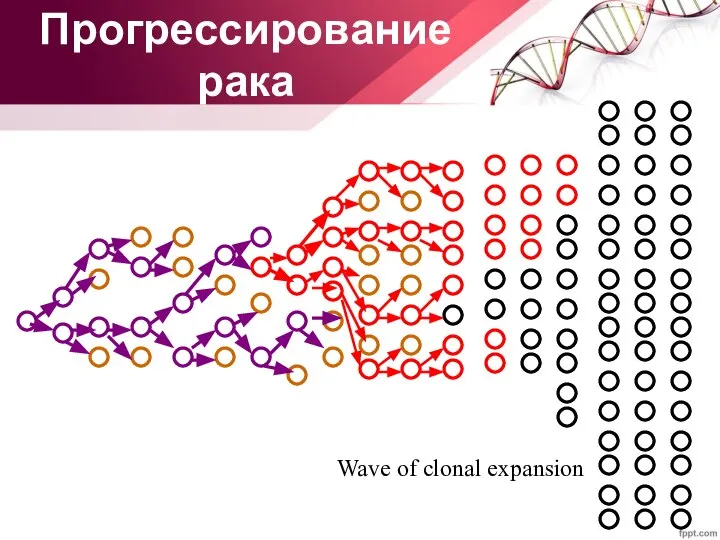
Wave of clonal expansion
Прогрессирование
рака
Wave of clonal expansion
Прогрессирование
рака
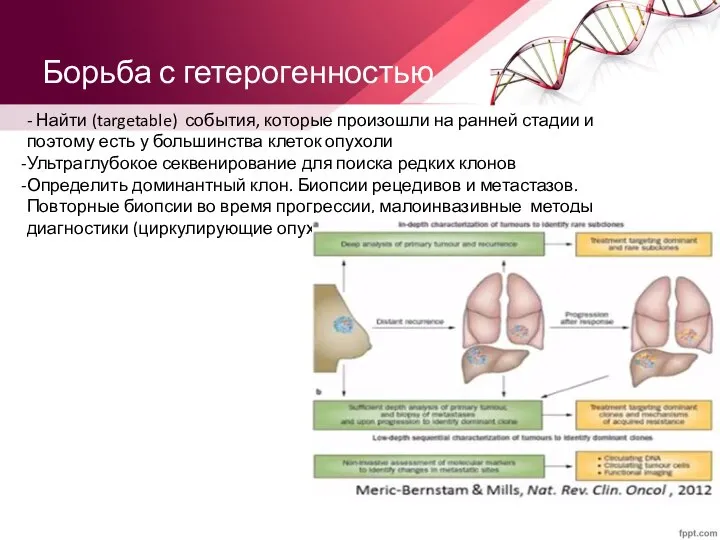
Борьба с гетерогенностью
- Найти (targetable) события, которые произошли на ранней стадии
Борьба с гетерогенностью
- Найти (targetable) события, которые произошли на ранней стадии

Мутации драйверы и мутации
пассажиры
Мутации драйверы и мутации
пассажиры

Возникающие генетические изменения в опухолевых клетках не всегда приводят к функциональным
Возникающие генетические изменения в опухолевых клетках не всегда приводят к функциональным

Генетические
нарушения
Делеции целых хромосомных районов, содержащих гены супрессоры опухолевого
Генетические
нарушения
Делеции целых хромосомных районов, содержащих гены супрессоры опухолевого

ЭПИГЕНЕТИЧЕСКИЕ
ФАКТОРЫ
наследственные и ненаследственные изменения в экспрессии конкретного гена без
ЭПИГЕНЕТИЧЕСКИЕ
ФАКТОРЫ
наследственные и ненаследственные изменения в экспрессии конкретного гена без
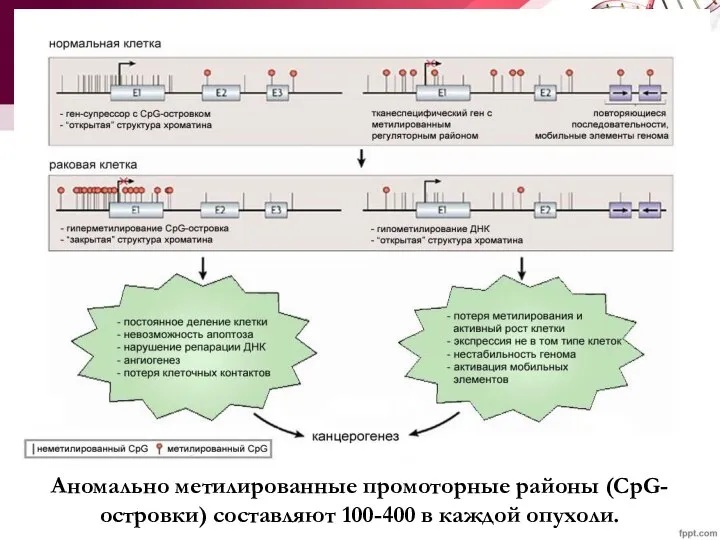
Аномально метилированные промоторные районы (CpG-островки) составляют 100-400 в каждой опухоли.
Аномально метилированные промоторные районы (CpG-островки) составляют 100-400 в каждой опухоли.

Метилирование ДНК является ценным биомаркером
для диагностики рака
1. Целый ряд
Метилирование ДНК является ценным биомаркером
для диагностики рака
1. Целый ряд

Частоты метилирования
генов-супрессоров
при раке почки
В опухолях метилирование VHL определено в
Частоты метилирования
генов-супрессоров
при раке почки
В опухолях метилирование VHL определено в

Методы анализа метилирования
1. Метилчувствительная ПЦР (NotI, EagI, SacII, HpaII, HhaI)
аналитическая
Методы анализа метилирования
1. Метилчувствительная ПЦР (NotI, EagI, SacII, HpaII, HhaI)
аналитическая
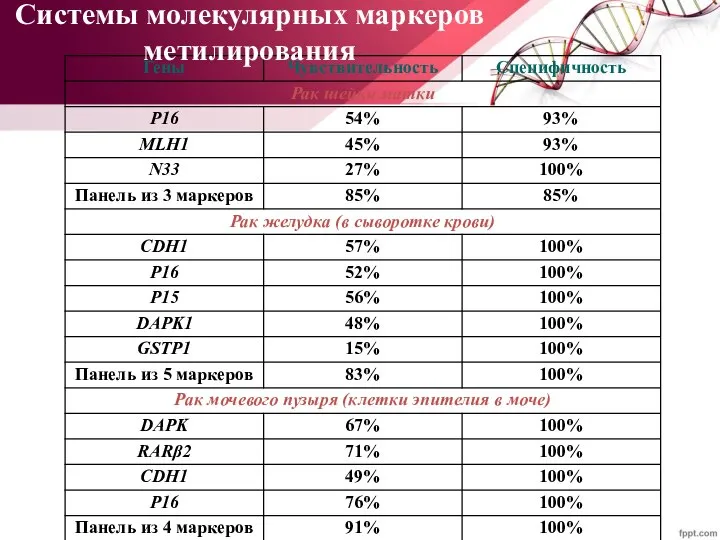
Системы молекулярных маркеров
метилирования
Системы молекулярных маркеров
метилирования

Метилирование промоторных районов генов RASSF и р16 достоверно чаще происходит
Метилирование промоторных районов генов RASSF и р16 достоверно чаще происходит

Метилирование позволяет предсказать поведение опухоли (эффективность терапии, метастазирование):
- метилирование WIT1
Метилирование позволяет предсказать поведение опухоли (эффективность терапии, метастазирование):
- метилирование WIT1

Метилирование ДНК, как диагностический маркер онкологического заболевания, имеет ряд преимуществ перед
Метилирование ДНК, как диагностический маркер онкологического заболевания, имеет ряд преимуществ перед

Молекулярные маркеры, определяемые на ранних стадиях канцерогенеза.
Молекулярные маркеры, определяемые на ранних стадиях канцерогенеза.

Метилирование позволяет более точно классифицировать типы и подтипы опухолей:
Метилирование позволяет более точно классифицировать типы и подтипы опухолей:
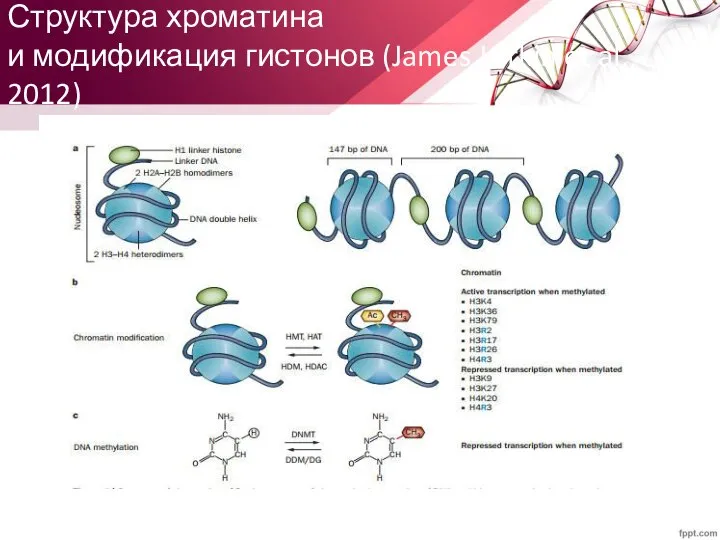
Структура хроматина
и модификация гистонов (James Larkin et al., 2012)
Структура хроматина
и модификация гистонов (James Larkin et al., 2012)
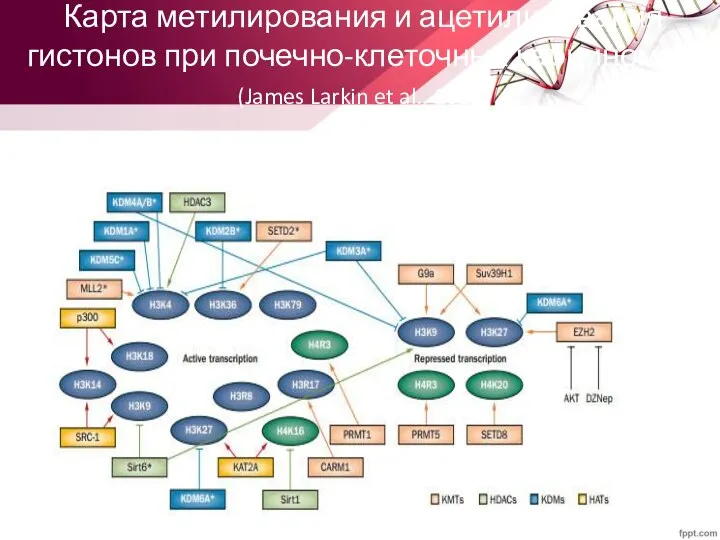
Карта метилирования и ацетилирования гистонов при почечно-клеточных карциномах
(James Larkin
Карта метилирования и ацетилирования гистонов при почечно-клеточных карциномах (James Larkin

РНК-интерференция: процесс подавления экспрессии гена на стадии транскрипции, трансляции, деаденилирования ил
РНК-интерференция: процесс подавления экспрессии гена на стадии транскрипции, трансляции, деаденилирования ил

МикроРНК - некодирующие РНК, состоящие из 18-24 нуклеотидов и регулирующие
МикроРНК - некодирующие РНК, состоящие из 18-24 нуклеотидов и регулирующие

Чиповые технологии
для высокопроизводительного генотипирования
Affymetrix
Agilent technologies
Genepix
Asper Biotech
Perkin Elmer
Illumina
Чиповые технологии
для высокопроизводительного генотипирования
Affymetrix
Agilent technologies
Genepix
Asper Biotech
Perkin Elmer
Illumina

Белок р53
Фактор апоптоза
p53 определяет повреждения ДНК
останавливает синтез ДНК до тех пор,
Белок р53
Фактор апоптоза
p53 определяет повреждения ДНК
останавливает синтез ДНК до тех пор,
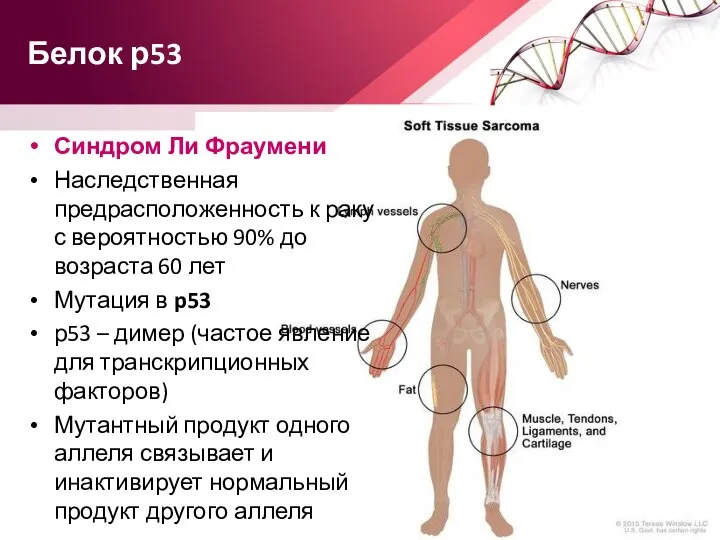
Белок р53
Синдром Ли Фраумени
Наследственная предрасположенность к раку с вероятностью 90% до
Белок р53
Синдром Ли Фраумени
Наследственная предрасположенность к раку с вероятностью 90% до
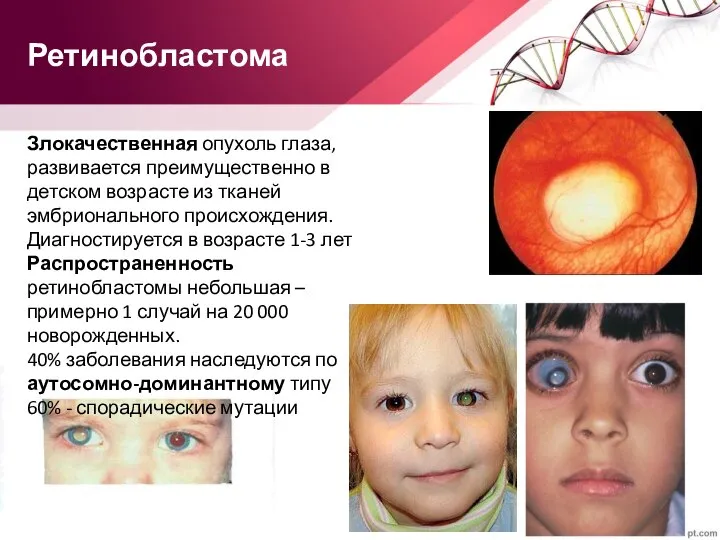
Ретинобластома
Злокачественная опухоль глаза, развивается преимущественно в детском возрасте из тканей эмбрионального
Ретинобластома
Злокачественная опухоль глаза, развивается преимущественно в детском возрасте из тканей эмбрионального
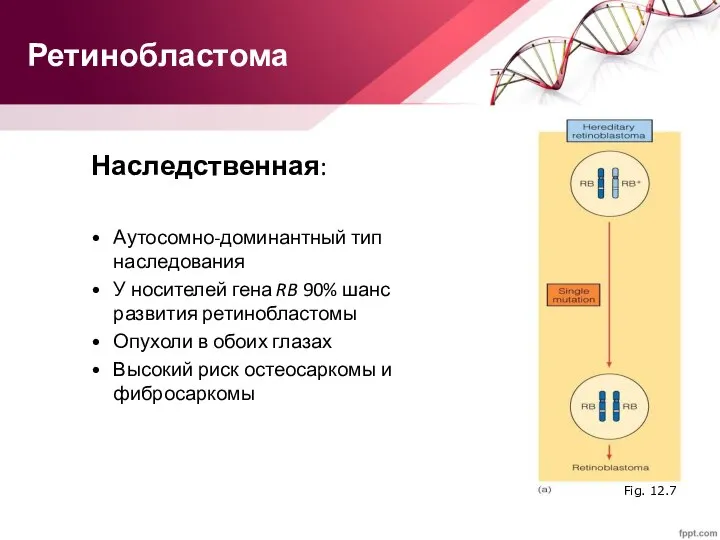
Ретинобластома
Наследственная:
Аутосомно-доминантный тип наследования
У носителей гена RB 90% шанс развития ретинобластомы
Опухоли в
Ретинобластома
Наследственная:
Аутосомно-доминантный тип наследования
У носителей гена RB 90% шанс развития ретинобластомы
Опухоли в
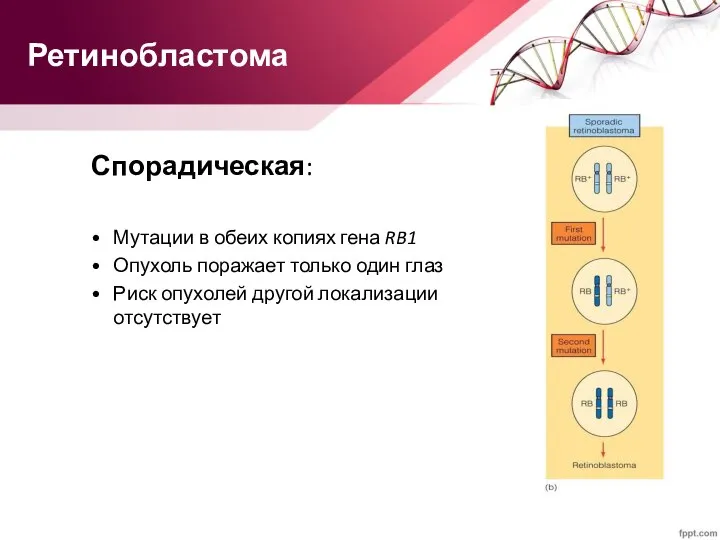
Ретинобластома
Спорадическая:
Мутации в обеих копиях гена RB1
Опухоль поражает только один глаз
Риск опухолей
Ретинобластома
Спорадическая:
Мутации в обеих копиях гена RB1
Опухоль поражает только один глаз
Риск опухолей
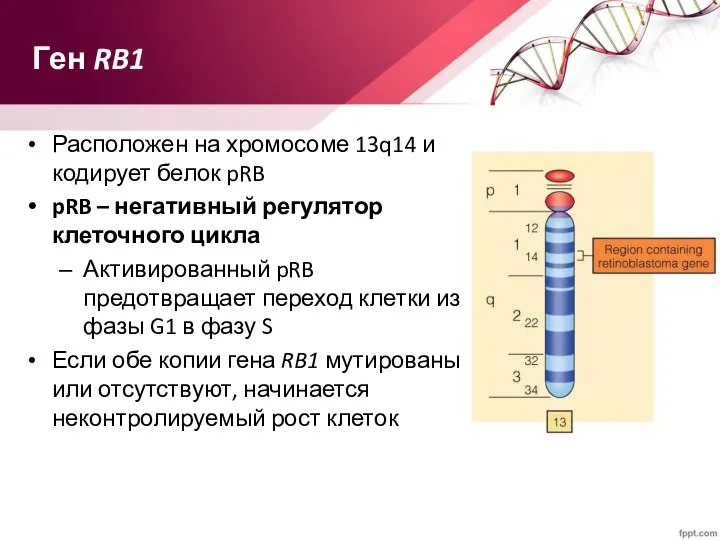
Ген RB1
Расположен на хромосоме 13q14 и кодирует белок pRB
pRB –
Ген RB1
Расположен на хромосоме 13q14 и кодирует белок pRB
pRB –
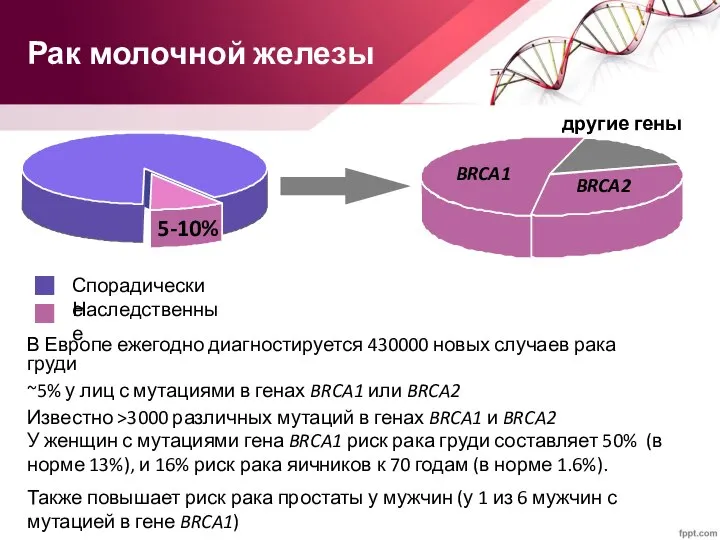
Рак молочной железы
В Европе ежегодно диагностируется 430000 новых случаев рака груди
~5%
Рак молочной железы
В Европе ежегодно диагностируется 430000 новых случаев рака груди
~5%

Критерии для постановки диагноза наследственного рака
молодой возраст возникновения онкологического заболевания;
Наследственный анамнез
Критерии для постановки диагноза наследственного рака
молодой возраст возникновения онкологического заболевания;
Наследственный анамнез
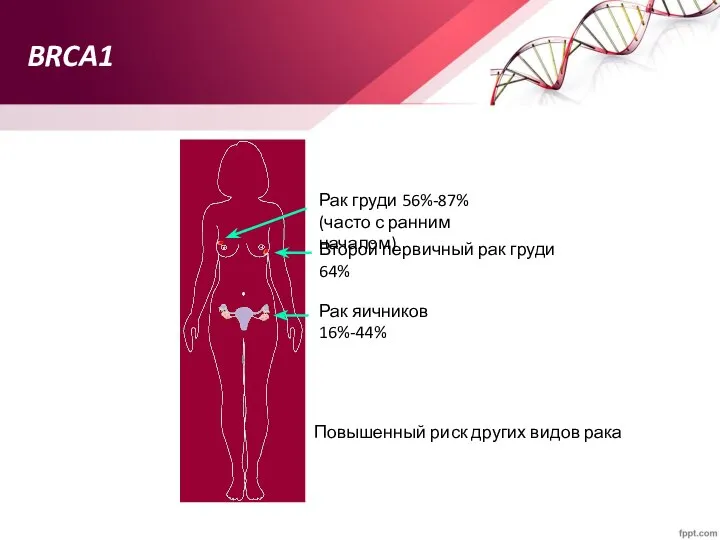
BRCA1
Повышенный риск других видов рака
Рак груди 56%-87%
(часто с ранним началом)
Второй
BRCA1
Повышенный риск других видов рака
Рак груди 56%-87%
(часто с ранним началом)
Второй
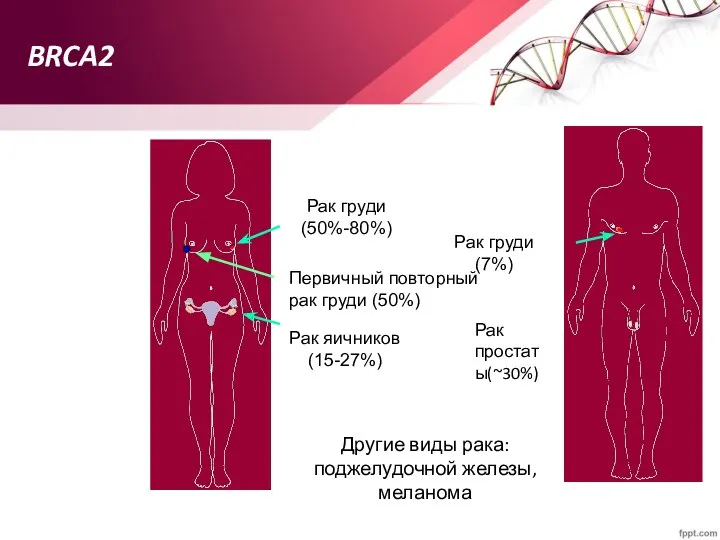
BRCA2
Другие виды рака: поджелудочной железы, меланома
Рак груди
(50%-80%)
Рак яичников
(15-27%)
Рак груди
(7%)
Первичный повторный рак
BRCA2
Другие виды рака: поджелудочной железы, меланома
Рак груди
(50%-80%)
Рак яичников
(15-27%)
Рак груди
(7%)
Первичный повторный рак

Экспрессия повышена в точке перехода из фазы G1 в фазу S
Экспрессия повышена в точке перехода из фазы G1 в фазу S

Скрининговые обследования:
Маммография/МРТ
Профилактическая мастэктомия
~90% снижение риска
Профилактическая двусторонняя сальпингооофория
~96-98% снижение риска рака яичников
~50%
Скрининговые обследования:
Маммография/МРТ
Профилактическая мастэктомия
~90% снижение риска
Профилактическая двусторонняя сальпингооофория
~96-98% снижение риска рака яичников
~50%


Показания:
2 родственников первой или второй степени с раком груди в возрасте
Показания:
2 родственников первой или второй степени с раком груди в возрасте
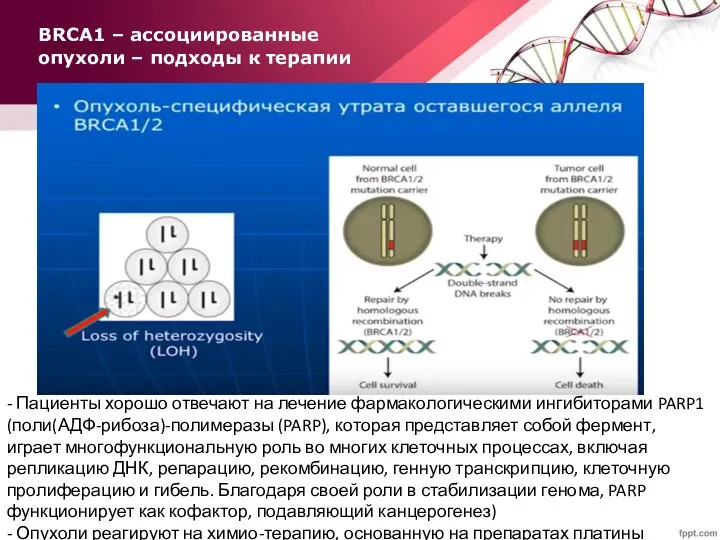
BRCA1 – ассоциированные опухоли – подходы к терапии
- Пациенты хорошо отвечают
BRCA1 – ассоциированные опухоли – подходы к терапии
- Пациенты хорошо отвечают
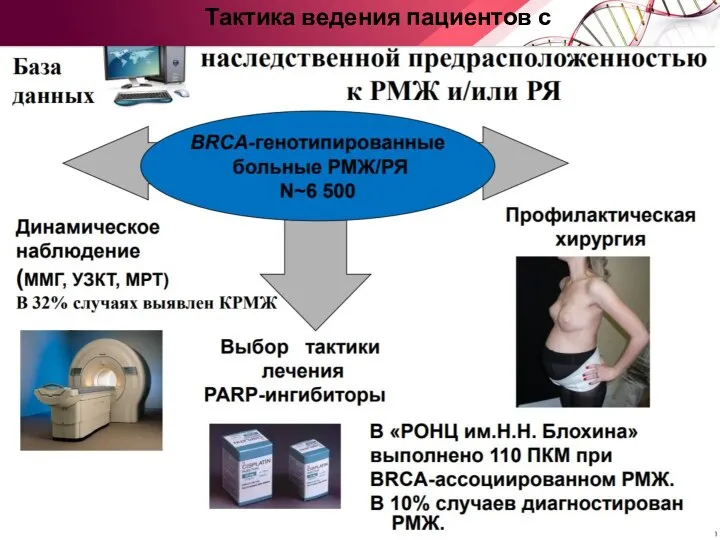
Тактика ведения пациентов с
Тактика ведения пациентов с

Наследственный рак предстательной железы
Наследственный рак предстательной железы
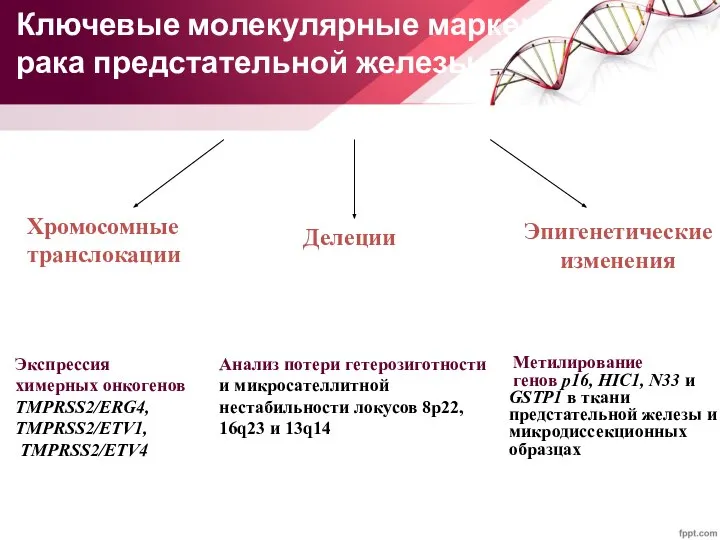
Ключевые молекулярные маркеры
рака предстательной железы
Эпигенетические
изменения
Метилирование
генов p16, HIC1, N33
Ключевые молекулярные маркеры
рака предстательной железы
Эпигенетические
изменения
Метилирование
генов p16, HIC1, N33

 Первичные иммунодефициты
Первичные иммунодефициты Системы регуляции жизнедеятельности
Системы регуляции жизнедеятельности Қан тобын анықтау
Қан тобын анықтау Концентрированные растворы
Концентрированные растворы Классификация методов психологического исследования
Классификация методов психологического исследования Лечебная физкультура при нарушениях осанки
Лечебная физкультура при нарушениях осанки Понятие о психике и ее эволюция
Понятие о психике и ее эволюция Роль медицинской сестры в профилактике инфекций, связанных с оказанием медицинской помощи
Роль медицинской сестры в профилактике инфекций, связанных с оказанием медицинской помощи Нарушения ритма сердца и проводимости
Нарушения ритма сердца и проводимости Врожденные формы автоматического поведения. (3)
Врожденные формы автоматического поведения. (3) ПЦР. Enzymatic amplification of beta-globin genomic sequences
ПЦР. Enzymatic amplification of beta-globin genomic sequences Жедел коронарлы синдромның асқынуы. Ауруханаға дейінгі этапта емдеу алгоритмы
Жедел коронарлы синдромның асқынуы. Ауруханаға дейінгі этапта емдеу алгоритмы Босанудан кейінгі акушерлік қан кету
Босанудан кейінгі акушерлік қан кету Профилактика гриппа и другие ОРВИ
Профилактика гриппа и другие ОРВИ Гигиена дыхания
Гигиена дыхания Слайд-лекция №3. Взаимодействие лекарственных средств
Слайд-лекция №3. Взаимодействие лекарственных средств Анафилактический шок у детей: этиология, патогенез, неотложная помощь, интенсивная терапия
Анафилактический шок у детей: этиология, патогенез, неотложная помощь, интенсивная терапия Профессия ветеринар
Профессия ветеринар Рациональная фармакотерапия острых респираторных вирусных инфекций
Рациональная фармакотерапия острых респираторных вирусных инфекций Что надо знать о коронавирусе
Что надо знать о коронавирусе Индивидуальный подход. Неврозы
Индивидуальный подход. Неврозы Нарушения водно-электролитного баланса, кровотечение
Нарушения водно-электролитного баланса, кровотечение Почечная колика
Почечная колика Язвенная болезнь желудка и двенадцатиперстной кишки
Язвенная болезнь желудка и двенадцатиперстной кишки Миофункциональный подход к профилактике и раннему лечению зубочелюстных аномалий
Миофункциональный подход к профилактике и раннему лечению зубочелюстных аномалий Денсаулық сақтаудағы тағам өнімдері
Денсаулық сақтаудағы тағам өнімдері Результаты мониторинга оказания медицинской помощи пациентам с онкологическими заболеваниями
Результаты мониторинга оказания медицинской помощи пациентам с онкологическими заболеваниями Современные лабораторные маркеры аутоиммунных заболеваний (СКВ, РА, склеродермия, дерматомиозит)
Современные лабораторные маркеры аутоиммунных заболеваний (СКВ, РА, склеродермия, дерматомиозит)