- Основные формы иммунного реагирования. Понятие об иммунитете. Виды иммунитета

Содержание
- 2. Антитела. Антителообразование. Киллинг. Иммунный фагоцитоз. Иммунная память и толерантность. Характеристика видов иммунитета. Особенности противовирусного, антипаразитарного и
- 3. Для гуморального иммунитета характерна выработка специфических антител (иммуноглобулинов). Антитела - специфические белки гамма- глобулиновой природы, образующиеся
- 4. Уникальность антител заключается в том, что они способны специфически взаимодействовать только с тем антигеном, который вызвал
- 5. Любая молекула антител имеет сходное строение ( Y- образную форму) и состоит из двух тяжелых (
- 6. Основные биологические характеристики антител. 1. Специфичность - способность взаимодействия с определенным (своим) антигеном (соответствие эпитопа антигена
- 7. Характеристика основных классов иммуноглобулинов. Ig G. Это основной класс иммуноглобулинов, защищающих организм от бактерий, токсинов и
- 8. IgM.. IgM- наиболее ранний класс антител, образующихся при первичном попадании антигена в организм. Наличие IgM- антител
- 9. IgA. Выделяют сывороточные IgA и секреторные IgA . Секреторные IgAs находятся в слюне, пищеварительных соках, секрете
- 11. С целью определения концентраций иммуноглобулинов отдельных классов применяют несколько методов, чаще используют метод радиальной иммунодиффузии в
- 12. Антитела класса IgM появляются в остром периоде заболевания и относительно быстро исчезают, антитела класса IgG выявляются
- 13. Любая молекула антител имеет сходное строение ( Y- образную форму) и состоит из двух тяжелых (
- 15. Антигенсвязывающий центр антител комплементарен эпитопу антигена по принципу “ключ - замок” и образован гипервариабельными областями L-
- 17. Выявлено пять классов тяжелых цепей- альфа ( с двумя подклассами), гамма ( с четырьмя подклассами), эксилон,
- 18. Антитела относятся преимущественно к γ-глобулиновой фракции белков плазмы крови.Поэтому антитела получили название иммуноглобулинов, и их обозначают
- 21. Антителопродукция в ответ на антигенный стимул имеет характерную динамику. Ее можно проследить на примере сывороточных Ig
- 22. В латентную фазу антителопродукция практически не изменяется и остается на базальном уровне. В этот период происходят
- 23. Во время логарифмической фазы наблюдается интенсивный прирост количества антигенспецифических В-лимфоцитов, что находит отражение в существенном нарастании
- 24. Повторный контакт иммунной системы с тем же антигеном ведет к формированию вторичного иммунного ответа. Его латентная
- 25. Это обусловлено подготовленностью иммунной системы к повторной встрече с антигеном за счет формирования иммунологической памяти :
- 26. Для развития гуморального иммунитета слизистых оболочек характерны те же процессы и динамика антителообразования. Однако в данном
- 27. Явление интенсивного антителообразования при повторном контакте с антигеном широко используется в практических целях, например вакцинопрофилактике. Для
- 28. Динамика и интенсивность антителообразования в значительной степени зависят от иммуногенности антигена: дозы, способа и кратности его
- 29. Роль антител в формировании иммунитета. Антитела имеют важное значение в формировании приобретенного постинфекционного и поствакцинального иммунитета.
- 30. 4. Антитела принимают участие в опсонизации бактерий, способствуя более эффективному фагоцитозу. 5. Антитела способствуют выведению из
- 31. Иммунный фагоцитоз Феномен иммунного фагоцитоза основан на поглощении фагоцитами антигенов, входящих в состав иммунных комплексов. При
- 32. Опосредованный клетками киллинг Иммунная система располагает независимым от системы комплемента способом уничтожения чужеродных клеток. Эта форма
- 33. Механизм клеточно-опосредованного киллинга достаточно универсален. Киллеры вырабатывают ряд веществ, которые вызывают нарушение целостности клеточной мембраны (или
- 34. Иммунологическая память. При повторной встрече с антигеном организм формирует более активную и быструю иммунную реакцию —
- 35. На сегодняшний день рассматривают два наиболее вероятных механизма формирования иммунологической памяти. Один из них предполагает длительное
- 36. Другой механизм предусматривает, что в процессе развития в организме продуктивного иммунного ответа часть антигенореактивных Т- или
- 37. Феномен иммунологической памяти широко используется в практике вакцинации людей для создания напряженного иммунитета и поддержания его
- 38. Иммунологическая толерантность — явление, противоположное иммунному ответу и иммунологической памяти. Проявляется она отсутствием специфического продуктивного иммунного
- 39. Иммунологическую толерантность вызывают антигены, которые получили название толерогены. Ими могут быть практически все вещества, однако наибольшей
- 40. Приобретенная толерантность может быть активной и пассивной. Активная толерантность создается путем введения в организм толерогена, который
- 41. Иммунологическая толерантность отличается специфичностью — она направлена к строго определенным антигенам. По степени распространенности различают поливалентную
- 42. Степень проявления иммунологической толерантности существенно зависит от ряда свойств макроорганизма и толерогена. Важное значение в индукции
- 43. Механизмы толерантности многообразны и до конца не расшифрованы. Известно, что ее основу составляют нормальные процессы регуляции
- 44. Феномен иммунологической толерантности имеет большое практическое значение. Он используется для решения многих важных проблем медицины, таких
- 45. Исторически иммунологическую толерантность рассматривают как защиту против аутоиммунных заболеваний. При нарушении толерантности к собственным антигенам могут
- 49. Основные механизмы отмены толерантности и развития аутоиммунных реакций 1. Изменения химической структуры аутоантигенов (например- изменение нормальной
- 50. Противобактериальный иммунитет основан на сочетанном воздействии на бактерии и их токсины факторов неспецифической резистентности (лизоцима, комплемента,
- 52. Противобактериальные антитела, в частности антитела к адгезинам бактерий, препятствуют прикреплению бактерий к тканям организма и, следовательно,
- 53. Антитела против токсинов бактерий участвуют в так называемом антитоксическом иммунитете. Антитоксические антитела, не влияя на колонизацию
- 54. Противовирусный иммунитет. Отличие противовирусного И. от других видов И. (против бактерий, простейших, грибков и т.д.) связано
- 57. Немаловажную роль в защите от вирусов играет воспалительная реакция, направленная на ограничение распространения вирусов в организме
- 59. Иммунитет при паразитарных болезнях. Различают частичный видовой и абсолютный (т.е. полную невосприимчивость человека к данному виду
- 60. Населению определенных зон тропической Африки свойственна полная невосприимчивость к возбудителю трехдневной малярииPlasmodium vivax вследствие утраты эритроцитами
- 61. Приобретенный И. при паразитарных заболеваниях обеспечивается теми же типами иммунного ответа, которые характерны для инфекций. Защитное
- 62. Нередко И. выражен только при наличии паразита в организме хозяина (нестерильный И., премуниция). Однако даже стерильный
- 64. Скачать презентацию

Антитела. Антителообразование.
Киллинг. Иммунный фагоцитоз.
Иммунная память и толерантность.
Характеристика видов
Антитела. Антителообразование.
Киллинг. Иммунный фагоцитоз.
Иммунная память и толерантность.
Характеристика видов

Для гуморального иммунитета характерна выработка специфических антител (иммуноглобулинов).
Антитела - специфические белки
Для гуморального иммунитета характерна выработка специфических антител (иммуноглобулинов).
Антитела - специфические белки

Уникальность антител заключается в том, что они способны специфически взаимодействовать только
Уникальность антител заключается в том, что они способны специфически взаимодействовать только

Любая молекула антител имеет сходное строение ( Y- образную форму) и
Любая молекула антител имеет сходное строение ( Y- образную форму) и

Основные биологические характеристики антител.
1. Специфичность - способность взаимодействия с определенным (своим)
Основные биологические характеристики антител.
1. Специфичность - способность взаимодействия с определенным (своим)

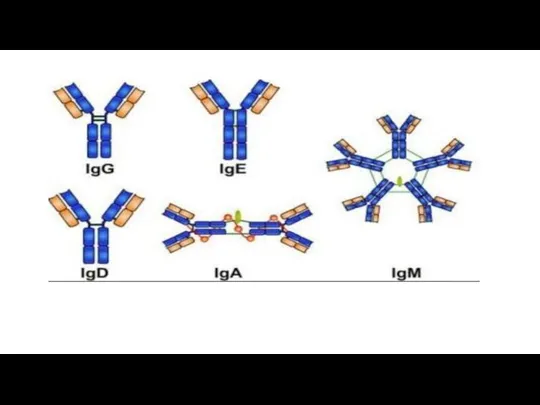
Характеристика основных классов иммуноглобулинов.
Ig G. Это основной класс иммуноглобулинов, защищающих организм
Характеристика основных классов иммуноглобулинов.
Ig G. Это основной класс иммуноглобулинов, защищающих организм

IgM.. IgM- наиболее ранний класс антител, образующихся при первичном попадании антигена
IgM.. IgM- наиболее ранний класс антител, образующихся при первичном попадании антигена

IgA. Выделяют сывороточные IgA и секреторные IgA . Секреторные IgAs находятся
IgA. Выделяют сывороточные IgA и секреторные IgA . Секреторные IgAs находятся

С целью определения концентраций иммуноглобулинов отдельных классов применяют несколько методов, чаще
С целью определения концентраций иммуноглобулинов отдельных классов применяют несколько методов, чаще

Антитела класса IgM появляются в остром периоде заболевания и относительно быстро
Антитела класса IgM появляются в остром периоде заболевания и относительно быстро

Любая молекула антител имеет сходное строение ( Y- образную форму) и
Любая молекула антител имеет сходное строение ( Y- образную форму) и
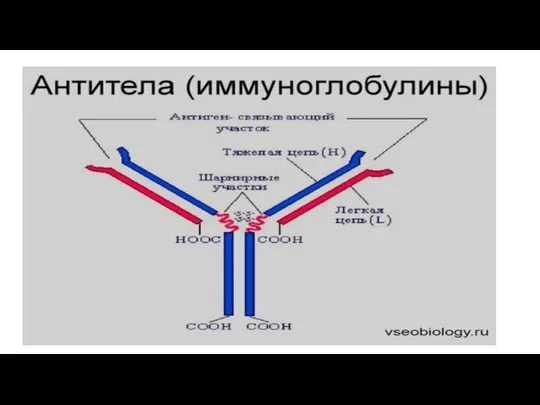

Антигенсвязывающий центр антител комплементарен эпитопу антигена по принципу “ключ - замок”
Антигенсвязывающий центр антител комплементарен эпитопу антигена по принципу “ключ - замок”


Выявлено пять классов тяжелых цепей- альфа ( с двумя подклассами), гамма
Выявлено пять классов тяжелых цепей- альфа ( с двумя подклассами), гамма

Антитела относятся преимущественно к γ-глобулиновой фракции белков плазмы крови.Поэтому антитела получили
Антитела относятся преимущественно к γ-глобулиновой фракции белков плазмы крови.Поэтому антитела получили


Антителопродукция в ответ на антигенный стимул имеет характерную динамику. Ее можно
Антителопродукция в ответ на антигенный стимул имеет характерную динамику. Ее можно

В латентную фазу антителопродукция практически не изменяется и остается на базальном уровне. В
В латентную фазу антителопродукция практически не изменяется и остается на базальном уровне. В

Во время логарифмической фазы наблюдается интенсивный прирост количества антигенспецифических В-лимфоцитов, что находит отражение
Во время логарифмической фазы наблюдается интенсивный прирост количества антигенспецифических В-лимфоцитов, что находит отражение

Повторный контакт иммунной системы с тем же антигеном ведет к формированию вторичного
Повторный контакт иммунной системы с тем же антигеном ведет к формированию вторичного

Это обусловлено подготовленностью иммунной системы к повторной встрече с антигеном за
Это обусловлено подготовленностью иммунной системы к повторной встрече с антигеном за

Для развития гуморального иммунитета слизистых оболочек характерны те же процессы и
Для развития гуморального иммунитета слизистых оболочек характерны те же процессы и

Явление интенсивного антителообразования при повторном контакте с антигеном широко используется в
Явление интенсивного антителообразования при повторном контакте с антигеном широко используется в

Динамика и интенсивность антителообразования в значительной степени зависят от иммуногенности антигена:
Динамика и интенсивность антителообразования в значительной степени зависят от иммуногенности антигена:

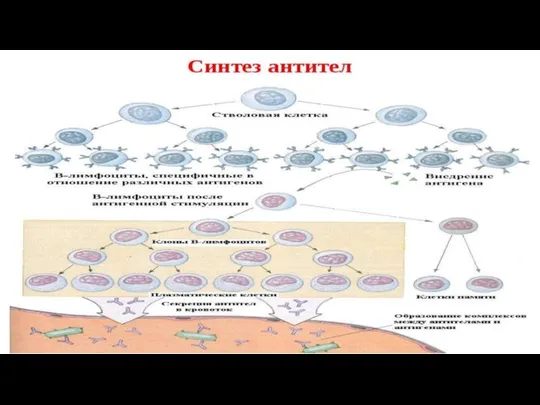
Роль антител в формировании иммунитета.
Антитела имеют важное значение в формировании приобретенного
Роль антител в формировании иммунитета.
Антитела имеют важное значение в формировании приобретенного

4. Антитела принимают участие в опсонизации бактерий, способствуя более эффективному фагоцитозу.
5.
4. Антитела принимают участие в опсонизации бактерий, способствуя более эффективному фагоцитозу.
5.

Иммунный фагоцитоз
Феномен иммунного фагоцитоза основан на поглощении фагоцитами антигенов, входящих в состав
Иммунный фагоцитоз
Феномен иммунного фагоцитоза основан на поглощении фагоцитами антигенов, входящих в состав

Опосредованный клетками киллинг
Иммунная система располагает независимым от системы комплемента способом
Опосредованный клетками киллинг
Иммунная система располагает независимым от системы комплемента способом

Механизм клеточно-опосредованного киллинга достаточно универсален.
Киллеры вырабатывают ряд веществ, которые вызывают
Механизм клеточно-опосредованного киллинга достаточно универсален.
Киллеры вырабатывают ряд веществ, которые вызывают

Иммунологическая память.
При повторной встрече с антигеном организм формирует более активную и
Иммунологическая память.
При повторной встрече с антигеном организм формирует более активную и

На сегодняшний день рассматривают два наиболее вероятных механизма формирования иммунологической памяти.
Один
На сегодняшний день рассматривают два наиболее вероятных механизма формирования иммунологической памяти.
Один

Другой механизм предусматривает, что в процессе развития в организме продуктивного иммунного
Другой механизм предусматривает, что в процессе развития в организме продуктивного иммунного

Феномен иммунологической памяти широко используется в практике вакцинации людей для создания
Феномен иммунологической памяти широко используется в практике вакцинации людей для создания

Иммунологическая толерантность — явление, противоположное иммунному ответу и иммунологической памяти. Проявляется она
Иммунологическая толерантность — явление, противоположное иммунному ответу и иммунологической памяти. Проявляется она

Иммунологическую толерантность вызывают антигены, которые получили название толерогены. Ими могут быть практически все
Иммунологическую толерантность вызывают антигены, которые получили название толерогены. Ими могут быть практически все

Приобретенная толерантность может быть активной и пассивной.
Активная толерантность создается путем введения в
Приобретенная толерантность может быть активной и пассивной.
Активная толерантность создается путем введения в

Иммунологическая толерантность отличается специфичностью — она направлена к строго определенным антигенам. По
Иммунологическая толерантность отличается специфичностью — она направлена к строго определенным антигенам. По

Степень проявления иммунологической толерантности существенно зависит от ряда свойств макроорганизма и толерогена.
Важное
Степень проявления иммунологической толерантности существенно зависит от ряда свойств макроорганизма и толерогена.
Важное

Механизмы толерантности многообразны и до конца не расшифрованы. Известно, что ее основу
Механизмы толерантности многообразны и до конца не расшифрованы. Известно, что ее основу

Феномен иммунологической толерантности имеет большое практическое значение. Он используется для решения многих
Феномен иммунологической толерантности имеет большое практическое значение. Он используется для решения многих

Исторически иммунологическую толерантность рассматривают как защиту против аутоиммунных заболеваний. При нарушении
Исторически иммунологическую толерантность рассматривают как защиту против аутоиммунных заболеваний. При нарушении




Основные механизмы отмены толерантности и развития аутоиммунных реакций
1. Изменения химической структуры
Основные механизмы отмены толерантности и развития аутоиммунных реакций
1. Изменения химической структуры

Противобактериальный иммунитет основан на сочетанном воздействии на бактерии и их токсины факторов неспецифической резистентности (лизоцима, комплемента,
Противобактериальный иммунитет основан на сочетанном воздействии на бактерии и их токсины факторов неспецифической резистентности (лизоцима, комплемента,


Противобактериальные антитела, в частности антитела к адгезинам бактерий, препятствуют прикреплению бактерий
Противобактериальные антитела, в частности антитела к адгезинам бактерий, препятствуют прикреплению бактерий

Антитела против токсинов бактерий участвуют в так называемом антитоксическом иммунитете. Антитоксические антитела,
Антитела против токсинов бактерий участвуют в так называемом антитоксическом иммунитете. Антитоксические антитела,

Противовирусный иммунитет. Отличие противовирусного И. от других видов И. (против бактерий,
Противовирусный иммунитет. Отличие противовирусного И. от других видов И. (против бактерий,

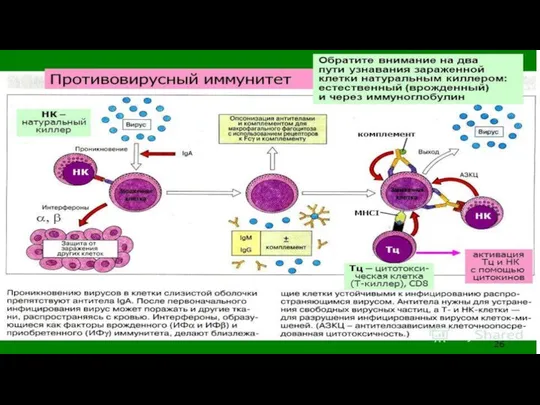

Немаловажную роль в защите от вирусов играет воспалительная реакция, направленная на ограничение
Немаловажную роль в защите от вирусов играет воспалительная реакция, направленная на ограничение


Иммунитет при паразитарных болезнях. Различают частичный видовой и абсолютный (т.е. полную невосприимчивость
Иммунитет при паразитарных болезнях. Различают частичный видовой и абсолютный (т.е. полную невосприимчивость

Населению определенных зон тропической Африки свойственна полная невосприимчивость к возбудителю трехдневной
Населению определенных зон тропической Африки свойственна полная невосприимчивость к возбудителю трехдневной

Приобретенный И. при паразитарных заболеваниях обеспечивается теми же типами иммунного ответа,
Приобретенный И. при паразитарных заболеваниях обеспечивается теми же типами иммунного ответа,

Нередко И. выражен только при наличии паразита в организме хозяина (нестерильный
Нередко И. выражен только при наличии паразита в организме хозяина (нестерильный
 Опухолевые стволовые клетки на примере рака молочной железы
Опухолевые стволовые клетки на примере рака молочной железы Сердечно-легочная реанимация
Сердечно-легочная реанимация Pathology of immune system
Pathology of immune system Роль патохарактерологических и микросоциальных факторов в развитии агрессивного и делинквентного поведения подростков
Роль патохарактерологических и микросоциальных факторов в развитии агрессивного и делинквентного поведения подростков Рентгенография ЖКТ с контрастом
Рентгенография ЖКТ с контрастом Псилоцибин - последствия наркотика, действие и признаки употребления
Псилоцибин - последствия наркотика, действие и признаки употребления Денсаулық сақтауға арналған ҚР бюджет жүйесі
Денсаулық сақтауға арналған ҚР бюджет жүйесі Дизартрия дегеніміз
Дизартрия дегеніміз Отчет по практике по педиатрии
Отчет по практике по педиатрии Хвороби дихальної системи
Хвороби дихальної системи Милдронат. История
Милдронат. История Физическая реабилитация при расстройствах обмена веществ (сахарный диабет)
Физическая реабилитация при расстройствах обмена веществ (сахарный диабет) Профилактика атеросклероза
Профилактика атеросклероза Для здорового баланса микрофлоры кишечника
Для здорового баланса микрофлоры кишечника Эффективность и хорошая переносимость слабительных средств
Эффективность и хорошая переносимость слабительных средств Обследование стоматологического больного. Асептика и антисептика в хирургической стоматологии
Обследование стоматологического больного. Асептика и антисептика в хирургической стоматологии Антиконвульсанты. Фармакология средств, корригирующих когнитивные функции. Фармакологическая регуляция церебрального кровотока
Антиконвульсанты. Фармакология средств, корригирующих когнитивные функции. Фармакологическая регуляция церебрального кровотока Водно-электролитные нарушения у хирургических больных и принципы инфузионной терапии
Водно-электролитные нарушения у хирургических больных и принципы инфузионной терапии Паренхиматозды мүшелердің жабық жарақаты
Паренхиматозды мүшелердің жабық жарақаты Магнитно-резонансная томография. МРТ. Режимы работы
Магнитно-резонансная томография. МРТ. Режимы работы Физиология возбудимых тканей и структур
Физиология возбудимых тканей и структур Психология и педагогика
Психология и педагогика Реактивтілік. Организм реактивтілігі
Реактивтілік. Организм реактивтілігі История развития психопатологии
История развития психопатологии Синдром гиперфибринолиза при шокогенной травме – насколько актуальна проблема
Синдром гиперфибринолиза при шокогенной травме – насколько актуальна проблема Мониторинг врожденных пороков развития
Мониторинг врожденных пороков развития Подростковый возраст 11 – 15 лет
Подростковый возраст 11 – 15 лет История развития психопатологии в России и зарубежных странах
История развития психопатологии в России и зарубежных странах