- Распространение возбуждения

Содержание
- 2. ПД –обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам.
- 3. Кодирование информации Согласно закону “все или ничего” амплитуда и длительность отдельных потенциалов действия постоянны Информация о
- 5. Кодирование информации Частота и количество в ряду зависит от интенсивности раздражения. Такой способ кодирования информации и
- 6. Из-за перезарядки мембраны во время генерации потенциала действия последний обладает способностью к самораспространению.
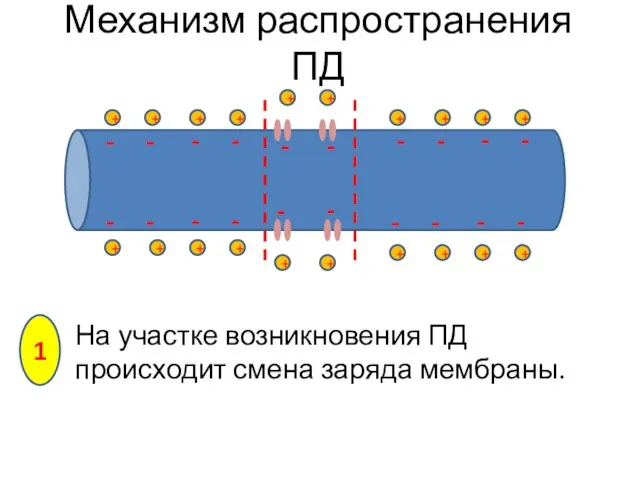
- 7. - - - - На участке возникновения ПД происходит смена заряда мембраны. Механизм распространения ПД -
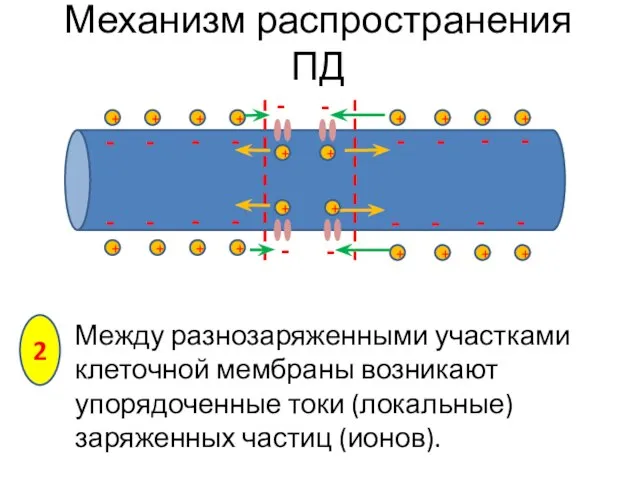
- 8. - - - - Между разнозаряженными участками клеточной мембраны возникают упорядоченные токи (локальные) заряженных частиц (ионов).
- 9. Локальные токи вызывают деполяризацию соседних невозбужденных участков мембраны. Механизм распространения ПД 3 - - - -
- 10. При достижении деполяризации КУД на невозбужденных участках мембраны формируется ПД. Механизм распространения ПД 4 - -
- 12. - - - - На участке перехвата Ранвье при возникновении ПД происходит смена заряда мембраны. Механизм
- 13. - - - - Между разнозаряженными перехватами Ранвье возникают упорядоченные токи (локальные) заряженных частиц (ионов). +
- 14. Локальные токи вызывают деполяризацию соседних невозбужденных перехватов Ранвье. 3 Механизм распространения ПД по миелиновому волокну -
- 15. При достижении деполяризации КУД на невозбужденных перехватах Ранвье формируется ПД. 4 Механизм распространения ПД по миелиновому
- 16. Наступающая после возбуждения в данном участке мембраны рефрактерность, обусловливает поступательное движение ПД.
- 17. Скорость распространения возбуждения по нервным волокнам разного типа: /—//—схема Опыта: а — установка, регистрирующая потенциалы нерва
- 19. ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ. Закон анатомической и физиологической непрерывности волокна. Любая травма волокна нарушает проводимость.
- 20. ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ. Закон двустороннего проведения возбуждения Однако, в целом организме по рефлекторной дуге
- 21. ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ. Закон изолированного проведения возбуждения в нервных стволах. Передача возбуждения на большие
- 22. ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ. Закон БЕЗДЕКРЕМЕНТНОГО проведения: ПД проходит весь путь от места раздражения до
- 23. СИНАПС Термин «синапс» (от греч. synapsis — соприкосновение, соединение) предложил в 1897 г. Чарлз Шеррингтон.
- 24. Синапс – специфическое место контакта (межклеточное мембранное соединение) возбудимых клеток, обеспечивающее передачу информации путем изменения потенциала
- 25. Электрические синапсы
- 26. Химические синапсы- информация передается химическим посредником — нейромедиатором.
- 27. ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ ПРЕСИНАПТИЧЕСКАЯ ЧАСТЬ ПОСТСИНАПТИЧЕСКАЯ ЧАСТЬ СИНАПТИЧЕСКАЯ ЩЕЛЬ
- 28. А/Х А Х ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ 1 + - + + - - + - +
- 29. Инактивация нейромедиатора. определяет кратковременность взаимодействия нейромедиатора с рецептором
- 30. Инактивация нейромедиатора. Диффузия в межклеточное пространство Ферментами (например, ацетилхолина — ацетилхолинэстеразой). Захват нейромедиатора. В большинстве синапсов
- 31. ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ Нейромедиаторы при связывании с ионотропными рецепторами приводят к возникновению ПСП. ПСП – колебания заряда
- 32. При возбуждении пресинаптической терминали ПД и секреции множества синаптических пузырьков регистрируются вызванные, или многоквантовые ПСП. Существуют
- 33. ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ Возбуждающие ПСП Тормозящие ПСП
- 34. Возбуждающие ПСП вызваны возрастанием проводимости мембраны для Na+. Они деполяризуют постсинаптическую мембрану, повышают возбудимость клетки, а
- 35. Активация н‑холинорецепторов и глутаматных (ионотропных) рецепторов приводит к возникновению возбуждающих ПСП. Пора (канал) этих рецепторов имеет
- 36. Тормозные ПСП вызваны повышением проводимости мембраны для K+ и Cl–. Они гиперполяризуют постсинаптическую мембрану, понижают возбудимость
- 37. ТПСП получил название постсинаптического торможения. Активация глициновых рецепторов и рецепторов ГАМК типа А приводит к возникновению
- 38. Быстрые (соматические) ПСП Медленные (вегетативные) ПСП
- 40. Центральные – в головном и спинном мозге, это межнейронные или нейрональные: аксосоматические аксодендритические аксоаксональные. Периферические –
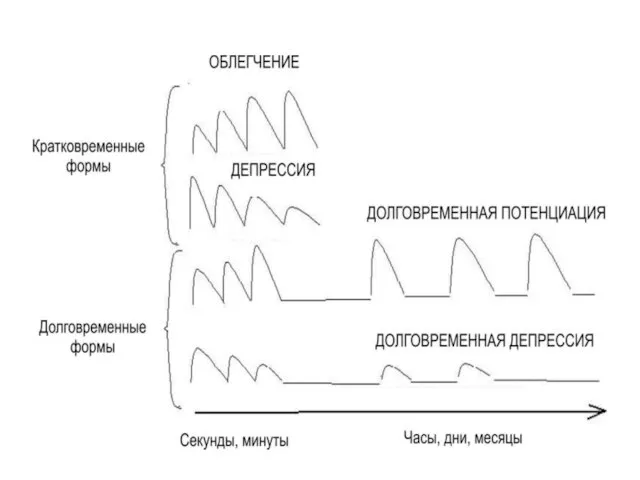
- 41. ПЛАСТИЧНОСТЬ СИНАПСОВ Пластичность может проявляться либо в увеличении (облегчении, потенциации), уменьшении (депрессии) эффективности синаптической передачи.
- 42. Выделяют кратковременные (длятся секунды и минуты) и долговременные (длятся часы, месяцы, годы) формы синаптической пластичности. Последние
- 45. Скачать презентацию
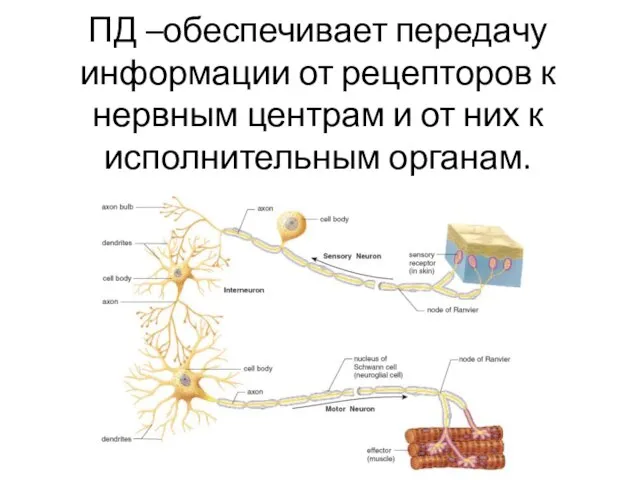
ПД –обеспечивает передачу информации от рецепторов к нервным центрам и от
ПД –обеспечивает передачу информации от рецепторов к нервным центрам и от

Кодирование информации
Согласно закону “все или ничего” амплитуда и длительность отдельных потенциалов
Кодирование информации
Согласно закону “все или ничего” амплитуда и длительность отдельных потенциалов


Кодирование информации
Частота и количество в ряду зависит от интенсивности раздражения.
Такой способ
Кодирование информации
Частота и количество в ряду зависит от интенсивности раздражения.
Такой способ
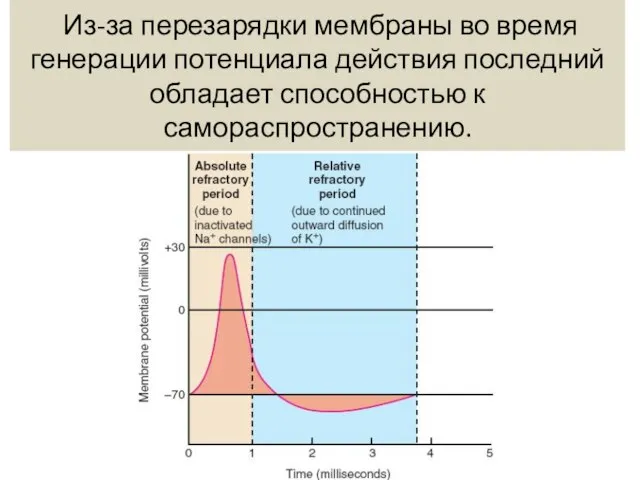
Из-за перезарядки мембраны во время генерации потенциала действия последний обладает
Из-за перезарядки мембраны во время генерации потенциала действия последний обладает
-
-
-
-
На участке возникновения ПД происходит смена заряда мембраны.
Механизм распространения ПД
-
-
-
-
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
1
+
-
-
-
-
На участке возникновения ПД происходит смена заряда мембраны.
Механизм распространения ПД
-
-
-
-
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
1
+
-
-
-
-
Между разнозаряженными участками клеточной мембраны возникают упорядоченные токи (локальные) заряженных частиц
-
-
-
-
Между разнозаряженными участками клеточной мембраны возникают упорядоченные токи (локальные) заряженных частиц
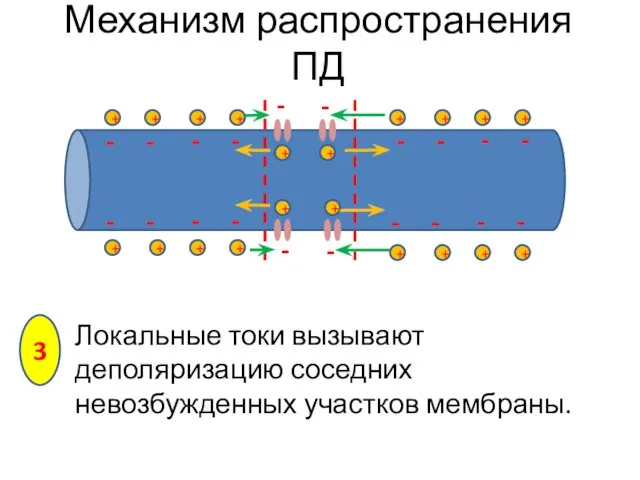
Локальные токи вызывают деполяризацию соседних невозбужденных участков мембраны.
Механизм распространения ПД
3
-
-
-
-
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Локальные токи вызывают деполяризацию соседних невозбужденных участков мембраны.
Механизм распространения ПД
3
-
-
-
-
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
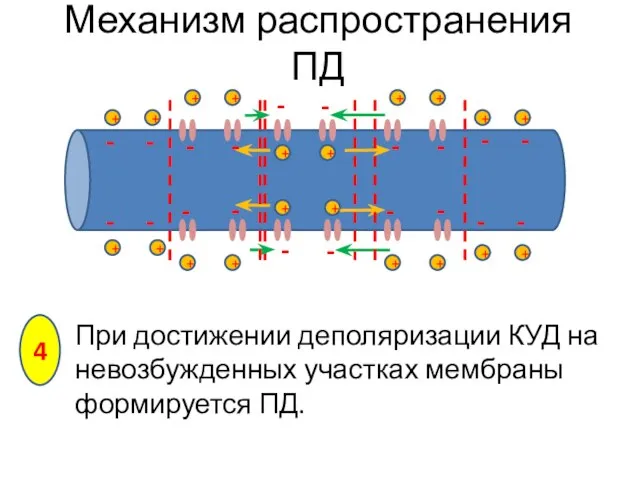
При достижении деполяризации КУД на невозбужденных участках мембраны формируется ПД.
Механизм распространения
При достижении деполяризации КУД на невозбужденных участках мембраны формируется ПД.
Механизм распространения
-
-
-
-
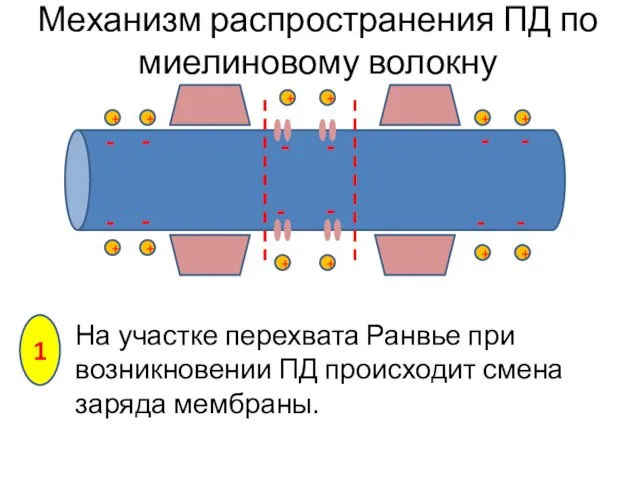
На участке перехвата Ранвье при возникновении ПД происходит смена заряда мембраны.
Механизм
-
-
-
-
На участке перехвата Ранвье при возникновении ПД происходит смена заряда мембраны.
Механизм
-
-
-
-
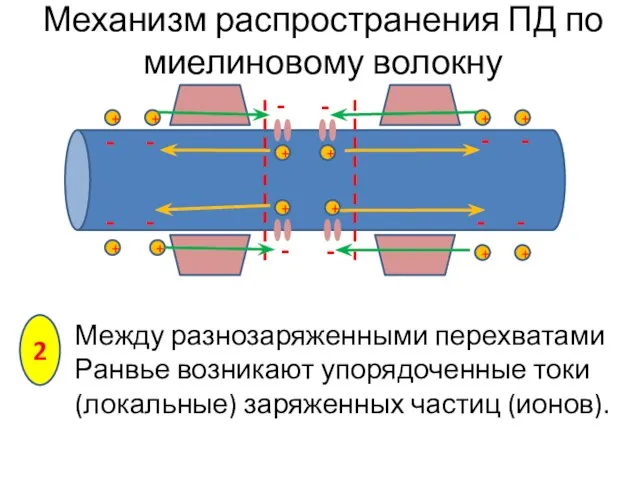
Между разнозаряженными перехватами Ранвье возникают упорядоченные токи (локальные) заряженных частиц (ионов).
+
+
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
2
Механизм
-
-
-
-
Между разнозаряженными перехватами Ранвье возникают упорядоченные токи (локальные) заряженных частиц (ионов).
+
+
+
+
+
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
2
Механизм
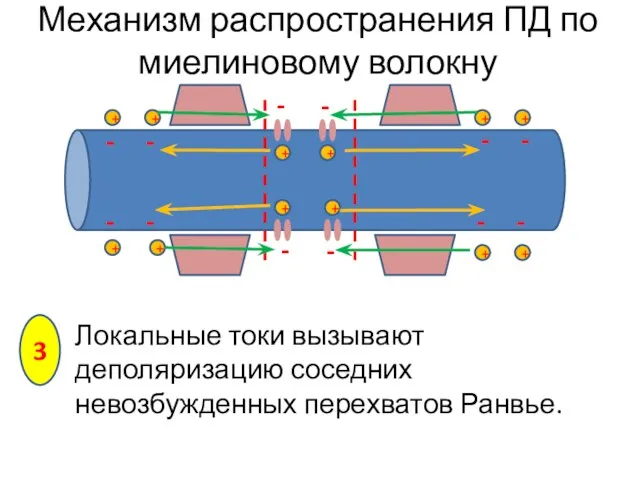
Локальные токи вызывают деполяризацию соседних невозбужденных перехватов Ранвье.
3
Механизм распространения ПД по
Локальные токи вызывают деполяризацию соседних невозбужденных перехватов Ранвье.
3
Механизм распространения ПД по
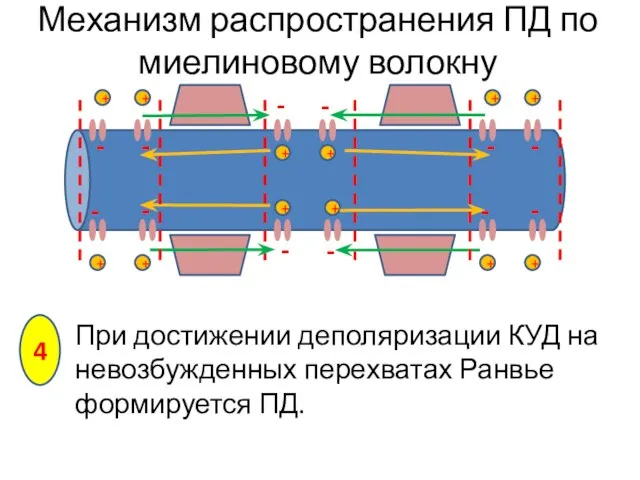
При достижении деполяризации КУД на невозбужденных перехватах Ранвье формируется ПД.
4
Механизм распространения
При достижении деполяризации КУД на невозбужденных перехватах Ранвье формируется ПД.
4
Механизм распространения
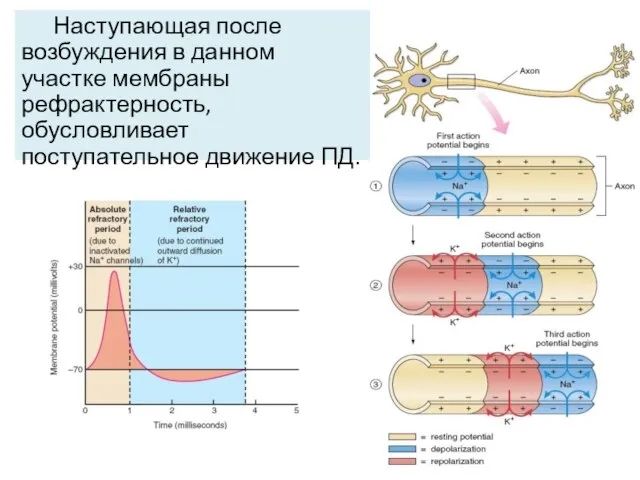
Наступающая после возбуждения в данном участке мембраны рефрактерность, обусловливает поступательное движение
Наступающая после возбуждения в данном участке мембраны рефрактерность, обусловливает поступательное движение
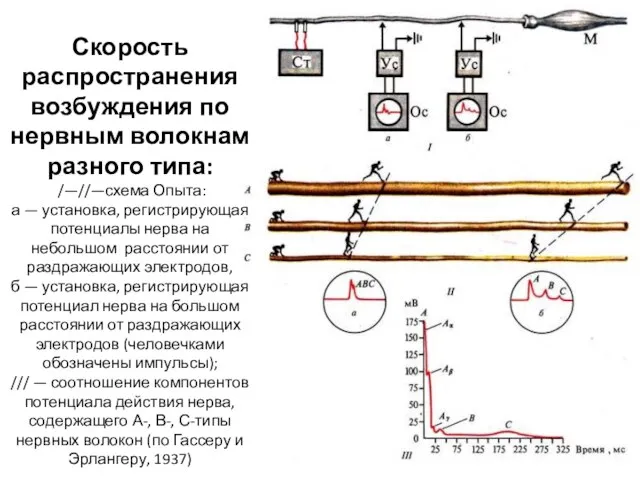
Скорость распространения возбуждения по нервным волокнам разного типа:
/—//—схема Опыта:
а
Скорость распространения возбуждения по нервным волокнам разного типа: /—//—схема Опыта: а

ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ.
Закон анатомической и физиологической непрерывности волокна.
Любая травма
ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ.
Закон анатомической и физиологической непрерывности волокна.
Любая травма

ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ.
Закон двустороннего проведения возбуждения
Однако, в целом организме
ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ.
Закон двустороннего проведения возбуждения
Однако, в целом организме

ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ.
Закон изолированного проведения возбуждения в нервных
ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ.
Закон изолированного проведения возбуждения в нервных

ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ.
Закон БЕЗДЕКРЕМЕНТНОГО проведения:
ПД проходит весь путь от
ЗАКОНЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ В НЕРВАХ.
Закон БЕЗДЕКРЕМЕНТНОГО проведения:
ПД проходит весь путь от

СИНАПС
Термин «синапс» (от греч. synapsis — соприкосновение, соединение) предложил в 1897 г.
СИНАПС
Термин «синапс» (от греч. synapsis — соприкосновение, соединение) предложил в 1897 г.

Синапс – специфическое место контакта
(межклеточное мембранное соединение)
возбудимых клеток, обеспечивающее
Синапс – специфическое место контакта
(межклеточное мембранное соединение)
возбудимых клеток, обеспечивающее
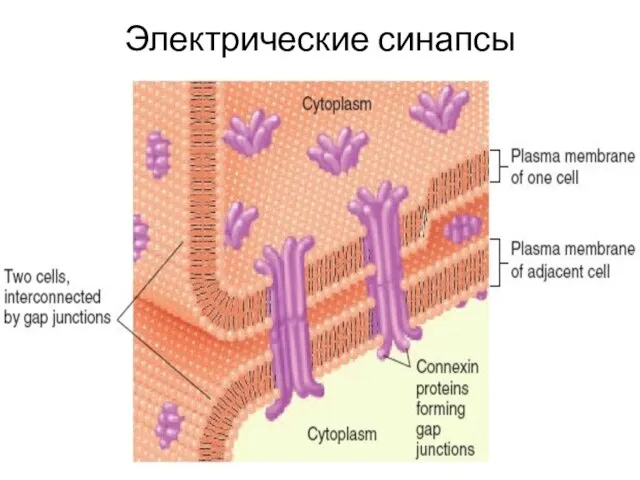
Электрические синапсы
Электрические синапсы
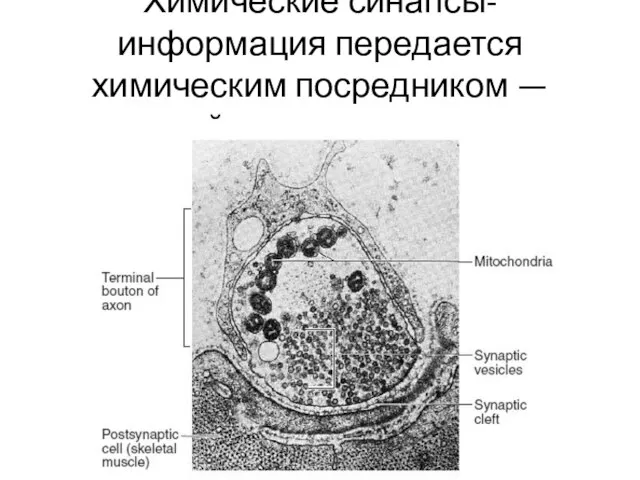
Химические синапсы- информация передается химическим посредником — нейромедиатором.
Химические синапсы- информация передается химическим посредником — нейромедиатором.
ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
ПРЕСИНАПТИЧЕСКАЯ ЧАСТЬ
ПОСТСИНАПТИЧЕСКАЯ ЧАСТЬ
СИНАПТИЧЕСКАЯ ЩЕЛЬ
ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
ПРЕСИНАПТИЧЕСКАЯ ЧАСТЬ
ПОСТСИНАПТИЧЕСКАЯ ЧАСТЬ
СИНАПТИЧЕСКАЯ ЩЕЛЬ
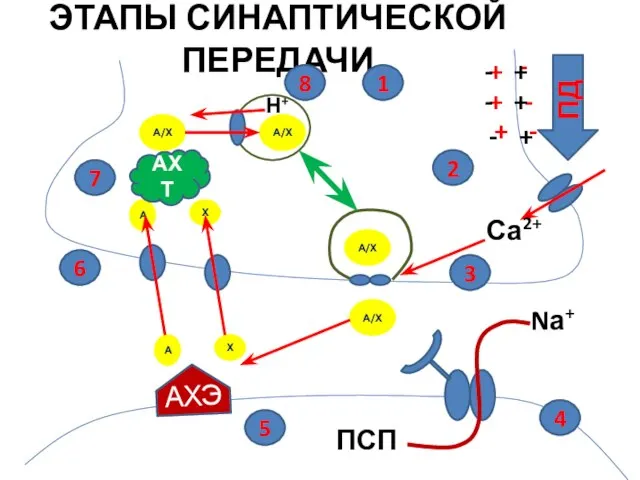
А/Х
А
Х
ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
1
+
-
+
+
-
-
+
-
+
-
+
-
ПД
2
8
3
4
6
5
7
Са2+
А/Х
А/Х
Nа+
ПСП
АХЭ
А
Х
АХТ
А/Х
Н+
А/Х
А
Х
ЭТАПЫ СИНАПТИЧЕСКОЙ ПЕРЕДАЧИ
1
+
-
+
+
-
-
+
-
+
-
+
-
ПД
2
8
3
4
6
5
7
Са2+
А/Х
А/Х
Nа+
ПСП
АХЭ
А
Х
АХТ
А/Х
Н+

Инактивация нейромедиатора.
определяет кратковременность взаимодействия нейромедиатора с рецептором
Инактивация нейромедиатора.
определяет кратковременность взаимодействия нейромедиатора с рецептором

Инактивация нейромедиатора.
Диффузия в межклеточное пространство
Ферментами (например, ацетилхолина — ацетилхолинэстеразой).
Захват нейромедиатора.
В
Инактивация нейромедиатора.
Диффузия в межклеточное пространство
Ферментами (например, ацетилхолина — ацетилхолинэстеразой).
Захват нейромедиатора.
В

ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ
Нейромедиаторы при связывании с ионотропными рецепторами приводят к возникновению ПСП.
ПСП
ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ
Нейромедиаторы при связывании с ионотропными рецепторами приводят к возникновению ПСП.
ПСП

При возбуждении пресинаптической терминали ПД и секреции множества синаптических пузырьков регистрируются
При возбуждении пресинаптической терминали ПД и секреции множества синаптических пузырьков регистрируются

ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ
Возбуждающие ПСП
Тормозящие ПСП
ПОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ
Возбуждающие ПСП
Тормозящие ПСП
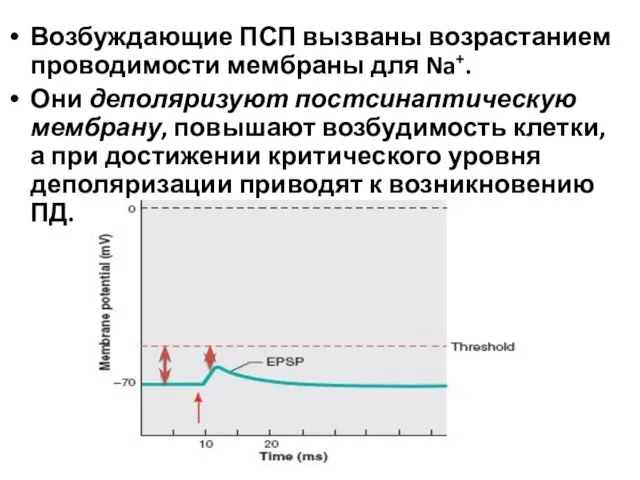
Возбуждающие ПСП вызваны возрастанием проводимости мембраны для Na+.
Они деполяризуют постсинаптическую
Возбуждающие ПСП вызваны возрастанием проводимости мембраны для Na+.
Они деполяризуют постсинаптическую

Активация н‑холинорецепторов и глутаматных (ионотропных) рецепторов приводит к возникновению возбуждающих ПСП.
Активация н‑холинорецепторов и глутаматных (ионотропных) рецепторов приводит к возникновению возбуждающих ПСП.
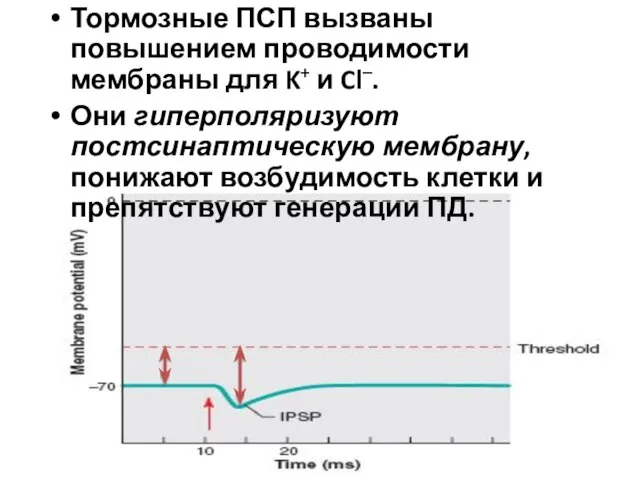
Тормозные ПСП вызваны повышением проводимости мембраны для K+ и Cl–.
Они
Тормозные ПСП вызваны повышением проводимости мембраны для K+ и Cl–.
Они

ТПСП получил название постсинаптического торможения.
Активация глициновых рецепторов и рецепторов ГАМК
ТПСП получил название постсинаптического торможения.
Активация глициновых рецепторов и рецепторов ГАМК

Быстрые (соматические) ПСП
Медленные (вегетативные) ПСП
Быстрые (соматические) ПСП
Медленные (вегетативные) ПСП

Центральные – в головном и спинном мозге, это межнейронные или нейрональные:
аксосоматические
аксодендритические
аксоаксональные.
Периферические
Центральные – в головном и спинном мозге, это межнейронные или нейрональные:
аксосоматические
аксодендритические
аксоаксональные.
Периферические

ПЛАСТИЧНОСТЬ СИНАПСОВ
Пластичность может проявляться либо в
увеличении (облегчении, потенциации),
уменьшении (депрессии)
ПЛАСТИЧНОСТЬ СИНАПСОВ
Пластичность может проявляться либо в
увеличении (облегчении, потенциации),
уменьшении (депрессии)

Выделяют
кратковременные (длятся секунды и минуты) и
долговременные (длятся часы, месяцы,
Выделяют
кратковременные (длятся секунды и минуты) и
долговременные (длятся часы, месяцы,
 Гонококковая инфекция у женщин
Гонококковая инфекция у женщин Атеросклероз
Атеросклероз Здоровое и сбалансированное питание
Здоровое и сбалансированное питание Хронические лейкозы (II часть)
Хронические лейкозы (II часть) Патофизиология нервной системы. Боль Моторная
Патофизиология нервной системы. Боль Моторная Заболевания новорожденных
Заболевания новорожденных Обеспечение охраны здоровья школьников в условиях распространения COVID-19
Обеспечение охраны здоровья школьников в условиях распространения COVID-19 Занятие физической культурой в послеродовом периоде
Занятие физической культурой в послеродовом периоде Первично-множественные опухоли у пациенток с раком яичников и раком молочной железы
Первично-множественные опухоли у пациенток с раком яичников и раком молочной железы Психиатриядағы емдеу әдістері
Психиатриядағы емдеу әдістері Методики определения предпочтительного типа будущей профессии
Методики определения предпочтительного типа будущей профессии Лямблиоз
Лямблиоз Патофизиология дисфункций системы пищеварения
Патофизиология дисфункций системы пищеварения Health is wealth
Health is wealth Захворювання органів сечовидільної системи та їх профілактика
Захворювання органів сечовидільної системи та їх профілактика Методы исследования блефароконъюнктивита грибково-демодекозной этиологии
Методы исследования блефароконъюнктивита грибково-демодекозной этиологии Острый медиастенит
Острый медиастенит Болезни крови и кроветворных органов у детей
Болезни крови и кроветворных органов у детей Лечение преэклампсии тяжелой степени в клинико-фармакологическом аспекте
Лечение преэклампсии тяжелой степени в клинико-фармакологическом аспекте Наркомании. Токсикомании
Наркомании. Токсикомании Диагностика повреждений опорно- двигательного аппарата
Диагностика повреждений опорно- двигательного аппарата Трикуспидальный стеноз
Трикуспидальный стеноз Генно-инженерные препараты в терапии ревматических заболеваний.pptx
Генно-инженерные препараты в терапии ревматических заболеваний.pptx Эффективное общение или как реагировать на неприемлемое поведение учеников
Эффективное общение или как реагировать на неприемлемое поведение учеников Количественные методы исследования осадка мочи
Количественные методы исследования осадка мочи Микоплазмы
Микоплазмы Школа диабета. Определение диабета
Школа диабета. Определение диабета Невынашивание беременности
Невынашивание беременности