- Тактильные ощущения и проприорецепция

Содержание
- 2. Механорецепторы
- 4. Схема расположения механорецепторов на неволосистых (А) и волосистых (Б) участках кожи. А Б С-волокна
- 5. Поверхностные кожные тактильные рецепторы
- 6. Поверхностные кожные тактильные рецепторы
- 7. Схема расположения механорецепторов на неволосистых (А) и волосистых (Б) участках кожи. А Б С-волокна
- 8. Тактильные рецепторы подкожной ткани
- 11. Пороговое различие двух точек для разных участков тела
- 12. Gating models of mechano-transduction channels. a, Stretch-activated ion channels open when forces (horizontal arrows) in the
- 13. Transduction channels (cyan) convert force into receptor currents, which then trigger action potentials by opening voltage-activated
- 14. Проприоцепция – ощущение положения и движения конечностей Мышечные веретена – медленно адаптирующиеся рецепторы. Чувствительны к изменению
- 15. Медиально-лемнисковая система Проведение тактильной и проприоцептивной информации
- 16. А. Схема возбудительных синаптических связей между тремя рецепторами и вставочными нейронами на следующих двух уровнях. Врезка
- 17. Латеральное торможение Три типа латерального торможения Локальные: Прямое (реципрокное) Возвратное Удаленное: 3) Дистальное
- 18. Эффект латерального торможения Тормозные вставочные нейроны (синие) на высших уровнях низводят частоту импульсации ниже уровня покоя
- 19. Удаленное: 3) Дистальное торможение Контроль сенсорного потока от периферических рецепторов со стороны мозговых структур
- 20. Влияние латерального торможения Торможение отсутствует на уровне рецептора, но присутствует на всех уровнях переключения
- 22. Карта тактильных проекций поверхности тела в области Бродмана 1, составленная по вызванным потенциалам Корреляция между областями
- 24. Рецептивные поля нейронов коры головного мозга Центральные нейроны имеют рецептивные поля. Каждая точка кожи представлена в
- 25. Рецептивные поля центральных нейронов неодинаковы для разных областей кожи.
- 26. Входы в соматосенсорную кору организованы в колонки по модальностям и субмодальностям Большинство клеток соматосенсорной системы реагируют
- 27. Mejsner Merkel Brodmann’s area 1
- 28. Интеграция сенсорной информации в нейронах коры головного мозга происходит за счет конвергенции и торможения при проведении
- 30. Фармакологическое угнетение нейронов коры в области Бродманна 2
- 31. Мимика обезьян Тонкая регуляция мимических мышц
- 32. Иллюзия Аристотеля Если скрестить два пальца и катать между ними горошину или какой-нибудь мелкий предмет, то
- 34. Мазохизм — склонность получать удовольствие, от насилия или мучения. Может быть чертой характера или девиацией в
- 36. Figure 1. Touch receptors in mammalian skin. Touch-sensitive afferents that innervate mammalian skin display morphological, functional,
- 37. Figure 3. Molecules that govern touch sensitivity in mammalian somatosensory neurons. Classes of ion channels that
- 39. Ardem Patapoutian) из Института Скриппса (США) обнаружила в нейронах, соединяющихся с клетками Меркеля, белок Piezo2, который
- 41. Скачать презентацию
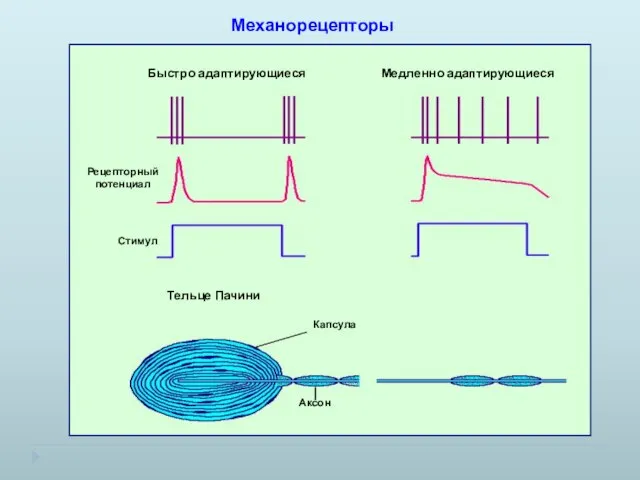
Механорецепторы
Механорецепторы
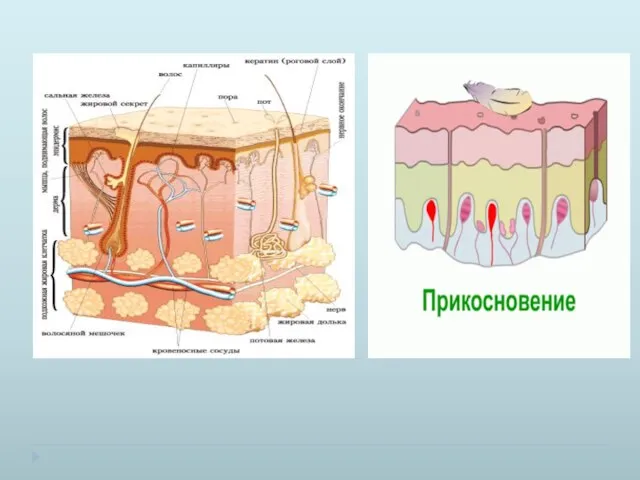
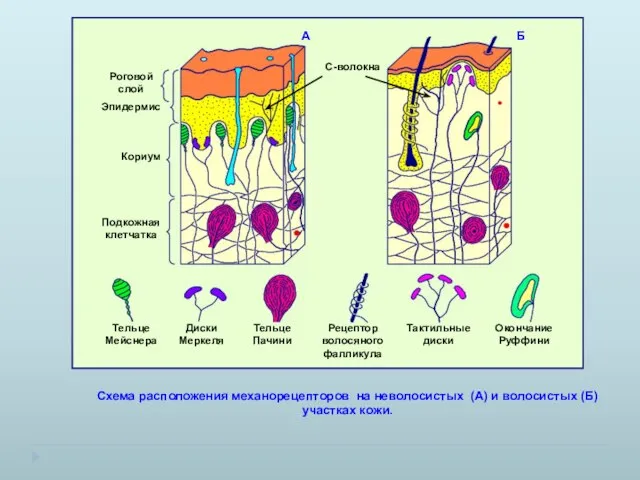
Схема расположения механорецепторов на неволосистых (А) и волосистых (Б) участках кожи.
А
Б
С-волокна
Схема расположения механорецепторов на неволосистых (А) и волосистых (Б) участках кожи.
А
Б
С-волокна

Поверхностные кожные тактильные рецепторы
Поверхностные кожные тактильные рецепторы

Поверхностные кожные тактильные рецепторы
Поверхностные кожные тактильные рецепторы
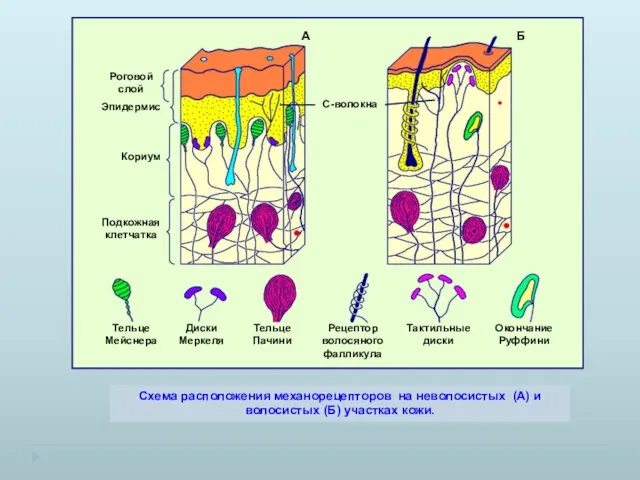
Схема расположения механорецепторов на неволосистых (А) и волосистых (Б) участках кожи.
А
Б
С-волокна
Схема расположения механорецепторов на неволосистых (А) и волосистых (Б) участках кожи.
А
Б
С-волокна
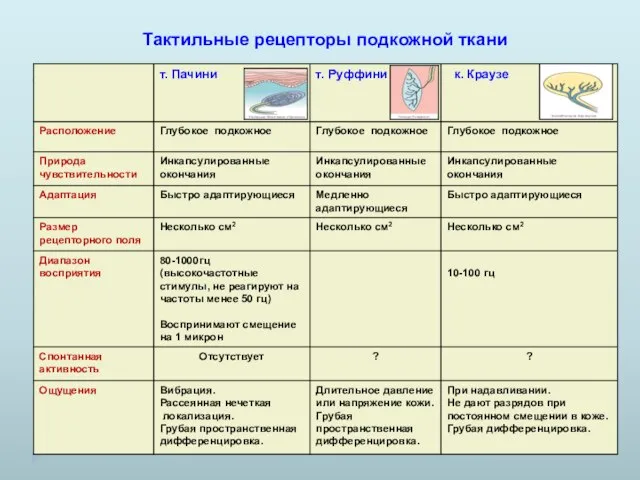
Тактильные рецепторы подкожной ткани
Тактильные рецепторы подкожной ткани
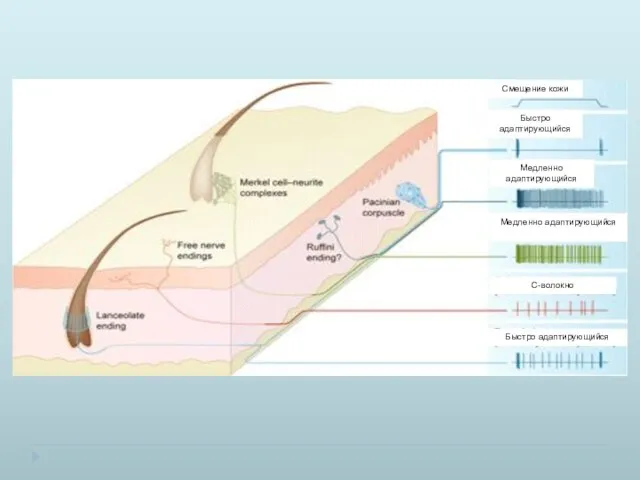
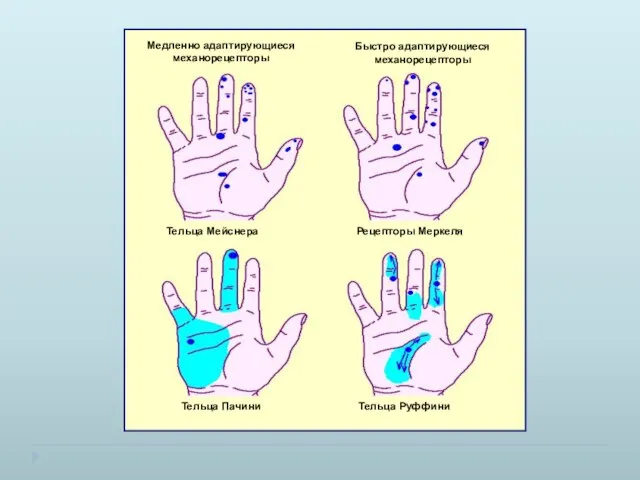
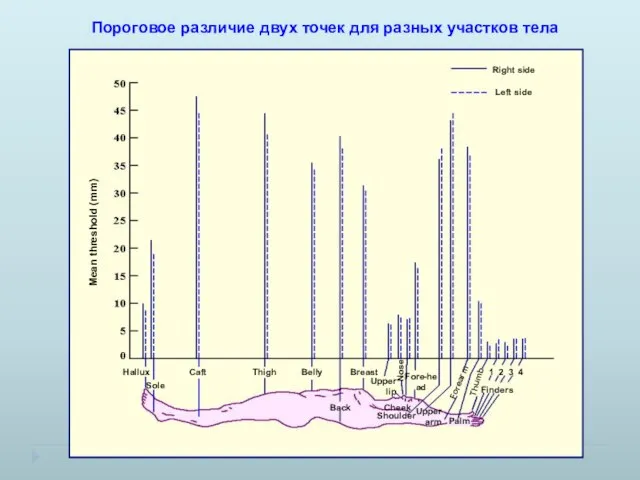
Пороговое различие двух точек для разных участков тела
Пороговое различие двух точек для разных участков тела
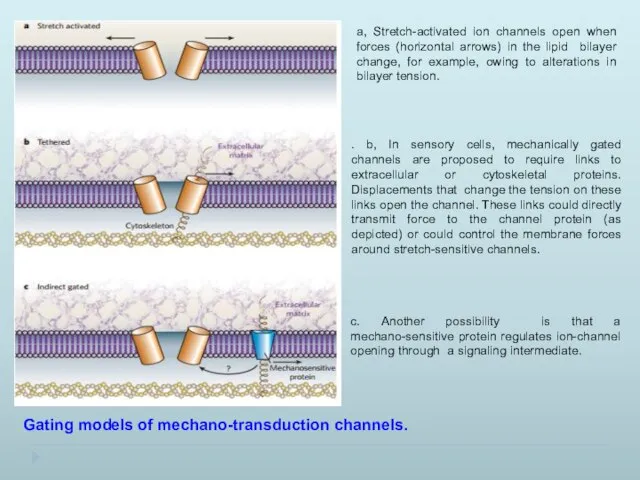
Gating models of mechano-transduction channels.
a, Stretch-activated ion channels open when
Gating models of mechano-transduction channels.
a, Stretch-activated ion channels open when
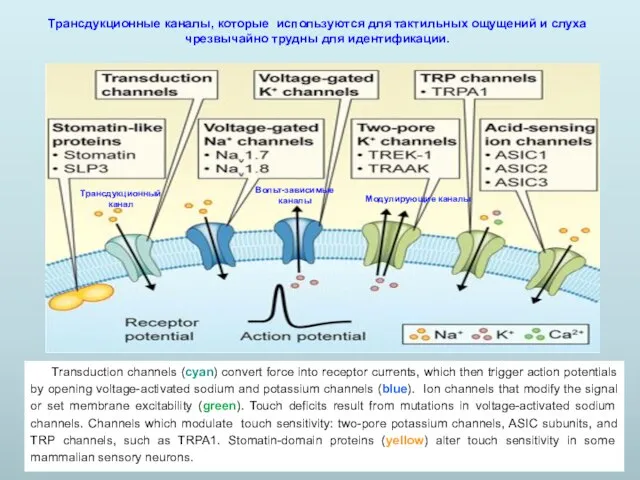
Transduction channels (cyan) convert force into receptor currents, which then trigger
Transduction channels (cyan) convert force into receptor currents, which then trigger
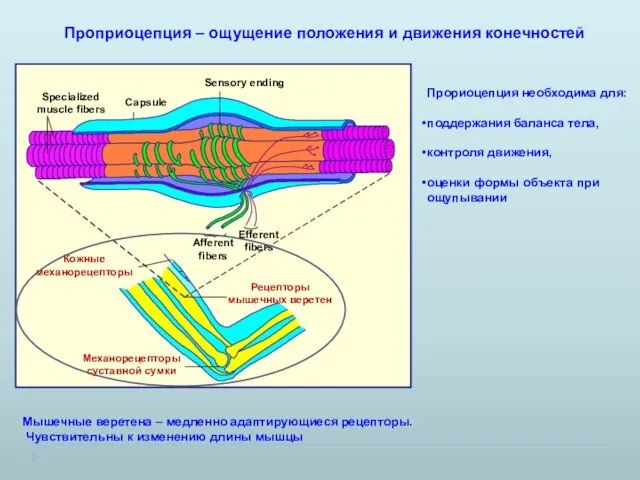
Проприоцепция – ощущение положения и движения конечностей
Мышечные веретена – медленно
Проприоцепция – ощущение положения и движения конечностей
Мышечные веретена – медленно
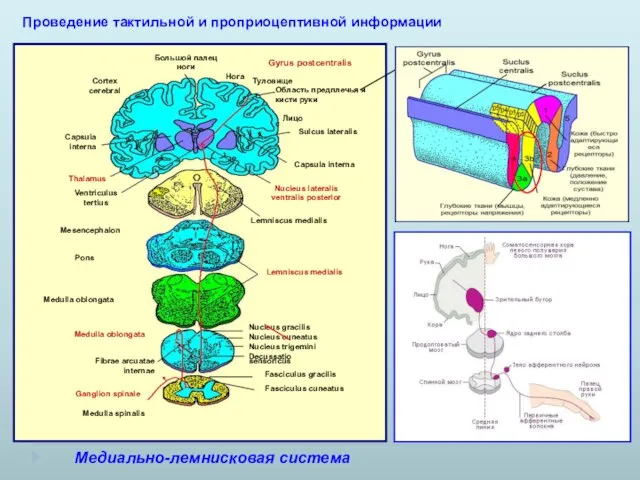
Медиально-лемнисковая система
Проведение тактильной и проприоцептивной информации
Медиально-лемнисковая система
Проведение тактильной и проприоцептивной информации
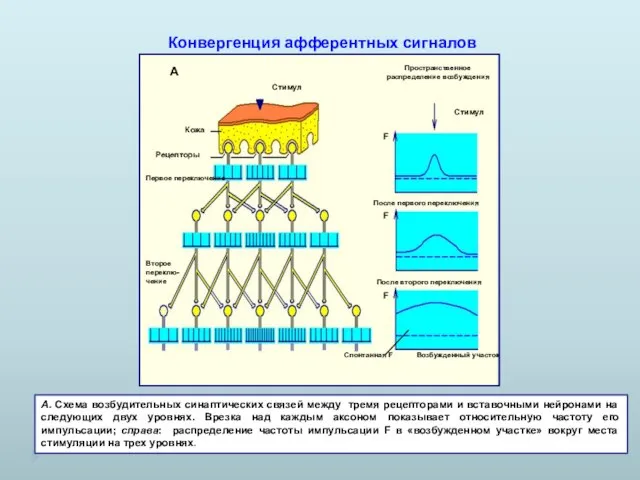
А. Схема возбудительных синаптических связей между тремя рецепторами и вставочными нейронами
А. Схема возбудительных синаптических связей между тремя рецепторами и вставочными нейронами
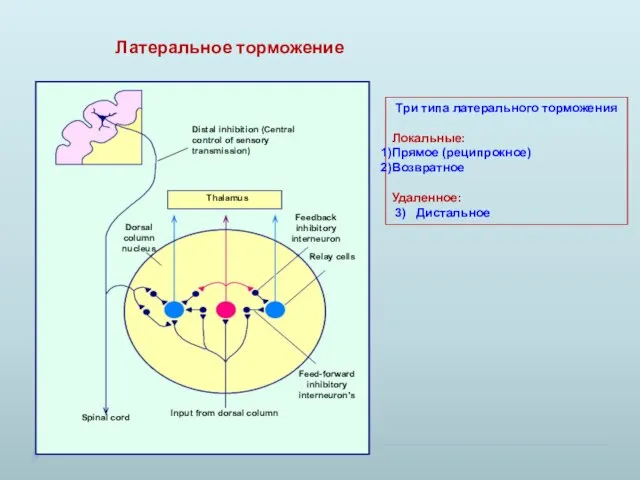
Латеральное торможение
Три типа латерального торможения
Локальные:
Прямое (реципрокное)
Возвратное
Удаленное:
3) Дистальное
Латеральное торможение
Три типа латерального торможения
Локальные:
Прямое (реципрокное)
Возвратное
Удаленное:
3) Дистальное
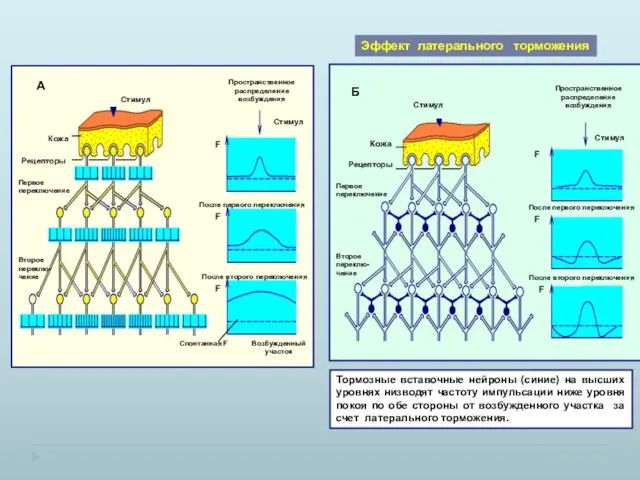
Эффект латерального торможения
Тормозные вставочные нейроны (синие) на высших уровнях низводят
Эффект латерального торможения
Тормозные вставочные нейроны (синие) на высших уровнях низводят
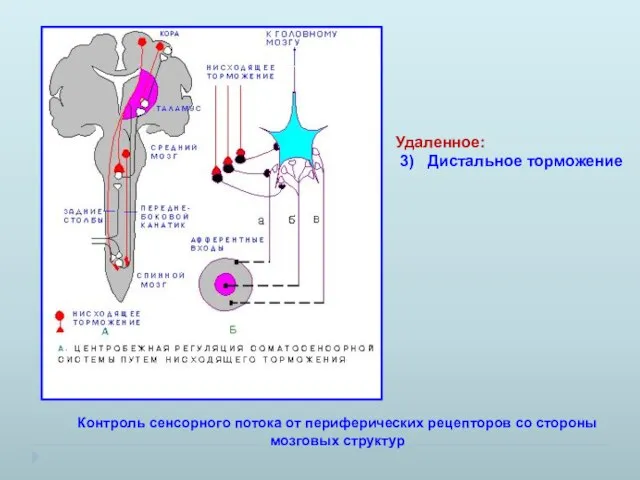
Удаленное:
3) Дистальное торможение
Контроль сенсорного потока от периферических рецепторов со стороны
Удаленное:
3) Дистальное торможение
Контроль сенсорного потока от периферических рецепторов со стороны
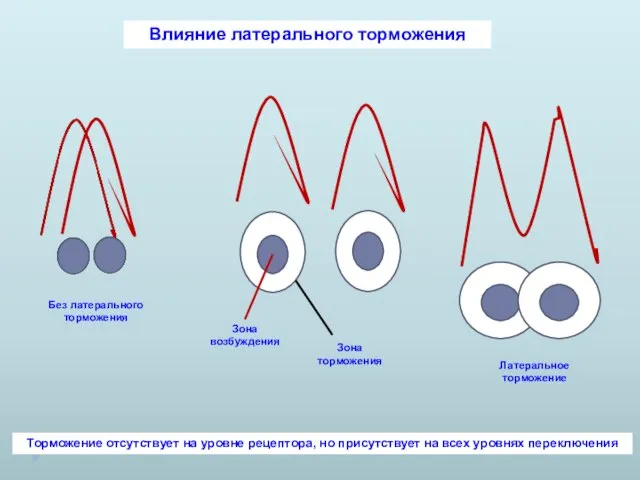
Влияние латерального торможения
Торможение отсутствует на уровне рецептора, но присутствует на всех
Влияние латерального торможения
Торможение отсутствует на уровне рецептора, но присутствует на всех

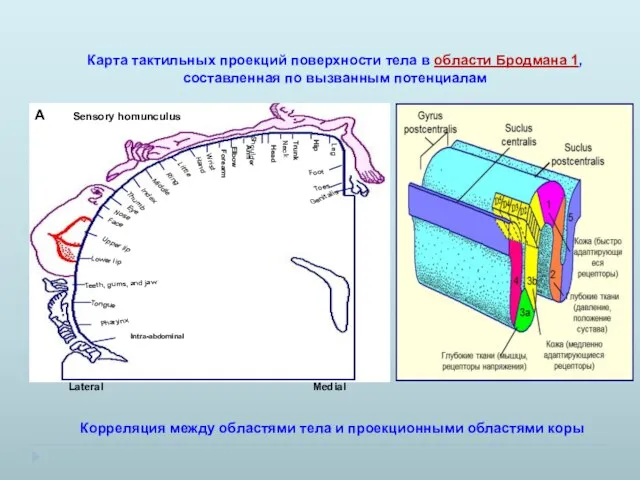
Карта тактильных проекций поверхности тела в области Бродмана 1, составленная по
Карта тактильных проекций поверхности тела в области Бродмана 1, составленная по
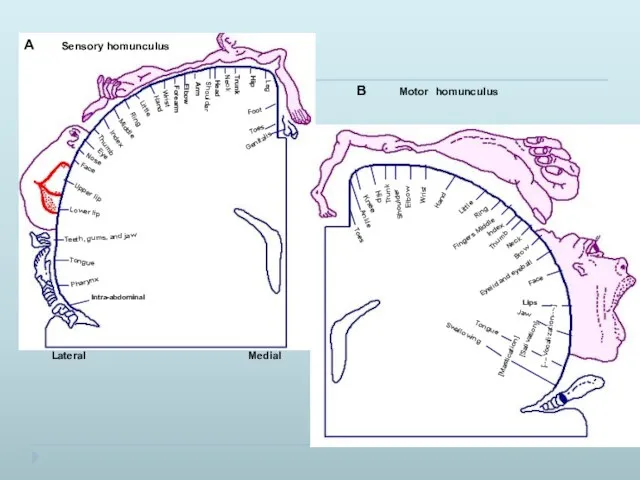

Рецептивные поля нейронов коры головного мозга
Центральные нейроны имеют рецептивные поля.
Каждая
Центральные нейроны имеют рецептивные поля.
Каждая
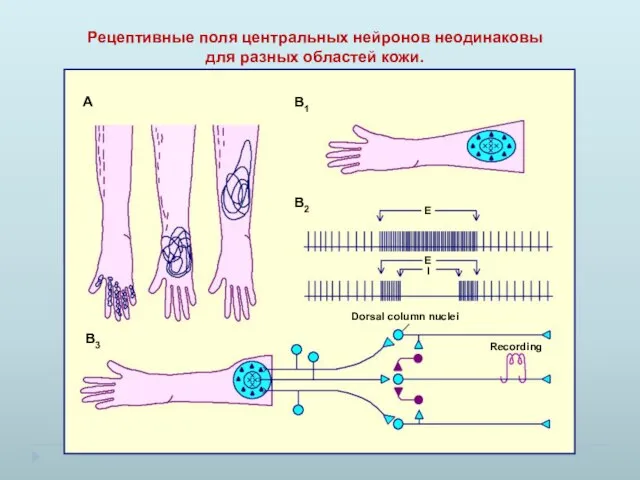
Рецептивные поля центральных нейронов неодинаковы для разных областей кожи.
Рецептивные поля центральных нейронов неодинаковы для разных областей кожи.
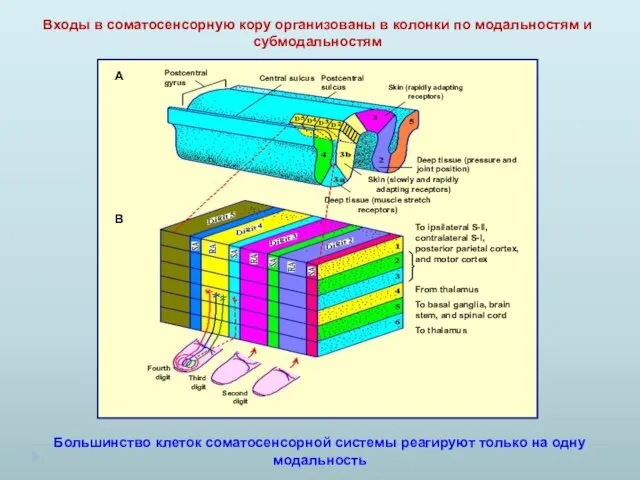
Входы в соматосенсорную кору организованы в колонки по модальностям и субмодальностям
Большинство
Входы в соматосенсорную кору организованы в колонки по модальностям и субмодальностям
Большинство

Mejsner
Merkel
Brodmann’s area 1
Mejsner
Merkel
Brodmann’s area 1

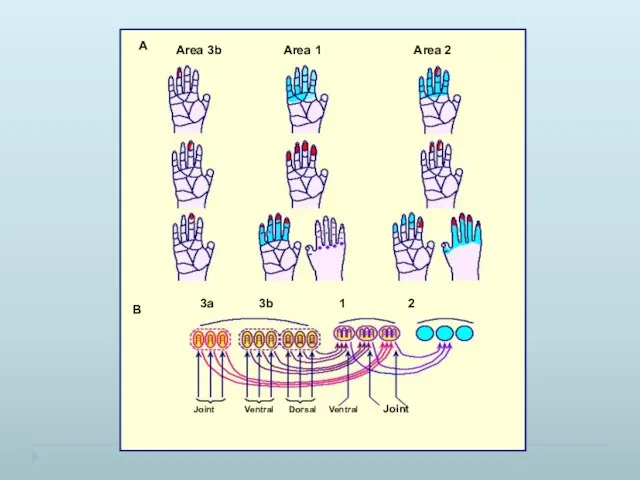
Интеграция сенсорной информации в нейронах коры головного мозга происходит за счет
Интеграция сенсорной информации в нейронах коры головного мозга происходит за счет
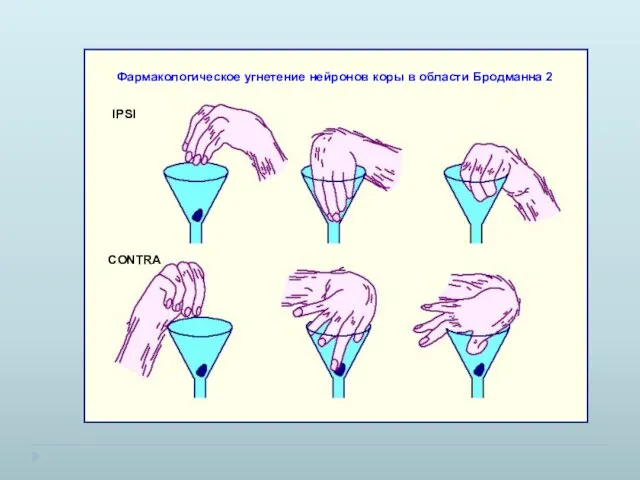
Фармакологическое угнетение нейронов коры в области Бродманна 2
Фармакологическое угнетение нейронов коры в области Бродманна 2
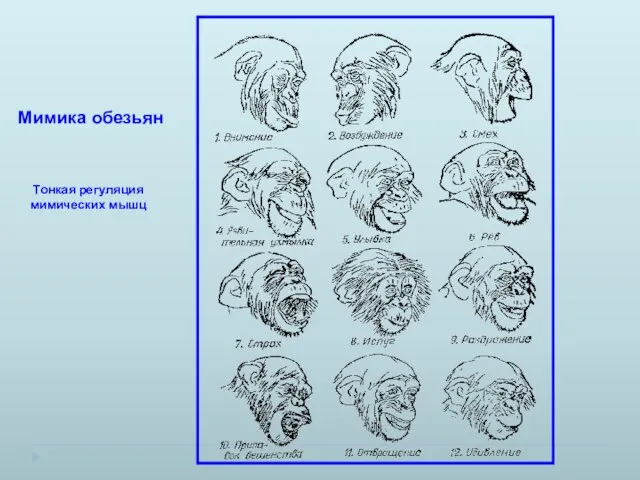
Мимика обезьян
Тонкая регуляция мимических мышц
Мимика обезьян
Тонкая регуляция мимических мышц

Иллюзия Аристотеля
Если скрестить два пальца и катать между ними горошину или
Иллюзия Аристотеля
Если скрестить два пальца и катать между ними горошину или


Мазохизм — склонность получать удовольствие, от насилия или мучения. Может быть
Мазохизм — склонность получать удовольствие, от насилия или мучения. Может быть
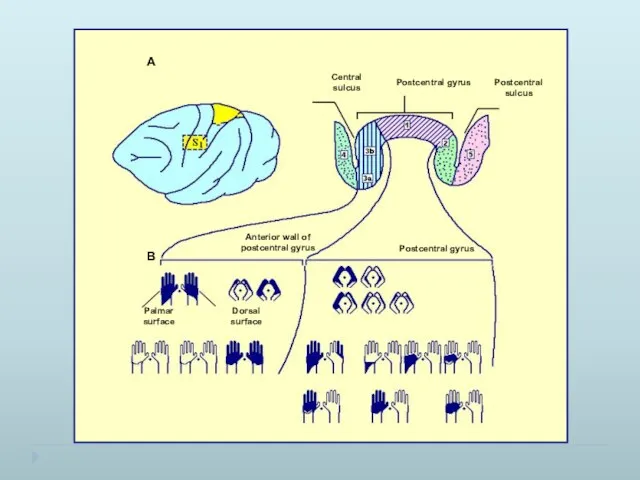
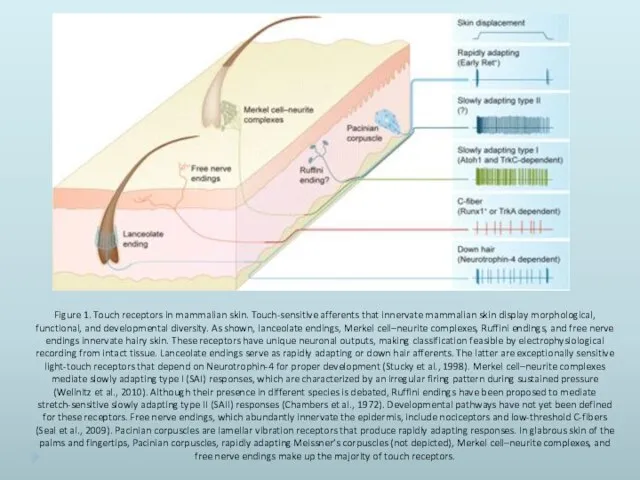
Figure 1. Touch receptors in mammalian skin. Touch-sensitive afferents that innervate
Figure 1. Touch receptors in mammalian skin. Touch-sensitive afferents that innervate
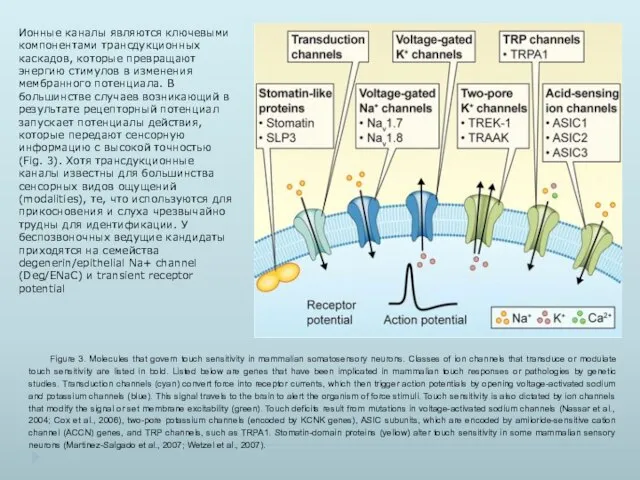
Figure 3. Molecules that govern touch sensitivity in mammalian somatosensory neurons.
Figure 3. Molecules that govern touch sensitivity in mammalian somatosensory neurons.

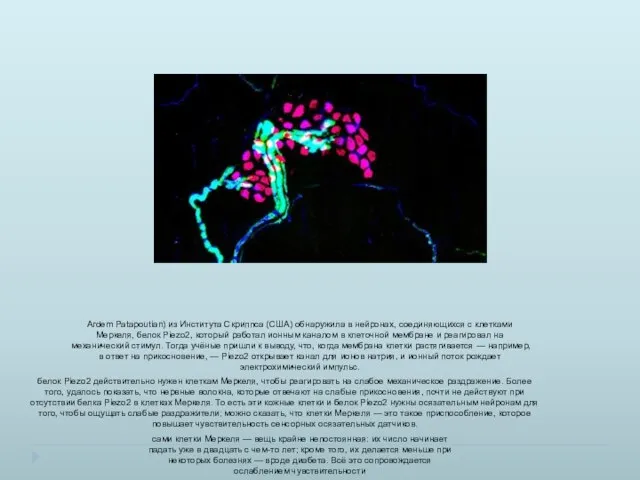
Ardem Patapoutian) из Института Скриппса (США) обнаружила в нейронах, соединяющихся с
Ardem Patapoutian) из Института Скриппса (США) обнаружила в нейронах, соединяющихся с
 Особенности работы участковой медсестры в детской поликлинике
Особенности работы участковой медсестры в детской поликлинике Эпилепсия
Эпилепсия Оценка профиля развития с использованием прикладной кинезиологии
Оценка профиля развития с использованием прикладной кинезиологии Аспирин
Аспирин Комбинированные гормональные методы
Комбинированные гормональные методы Современные методы помощи детям с РАС
Современные методы помощи детям с РАС Иондық сәулелену және оның ағзаға әсері
Иондық сәулелену және оның ағзаға әсері Кишечная Непроходимость
Кишечная Непроходимость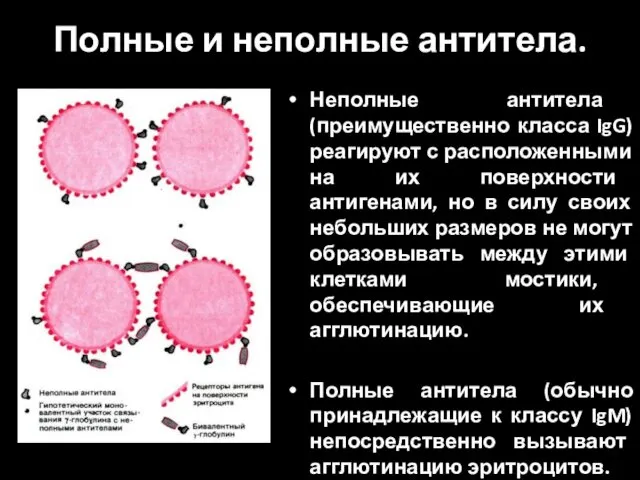 Полные и неполные антитела
Полные и неполные антитела 03-28.03 Шлифование и полирование
03-28.03 Шлифование и полирование Аномалии и самостоятельные заболевания языка. Хронические рецидивирующие трещины губ. Хейлиты
Аномалии и самостоятельные заболевания языка. Хронические рецидивирующие трещины губ. Хейлиты СПИД, возбудитель, течение, распространение
СПИД, возбудитель, течение, распространение Ауруды басқару бағдарламасына қатысатын сжж науқастардағы қауіп факторларының таралуын зерттеу
Ауруды басқару бағдарламасына қатысатын сжж науқастардағы қауіп факторларының таралуын зерттеу Лейкопоэзді қуатандыратың заттар. Препараттары - әсер ету механизмі, қолдану зәр айдағыштар
Лейкопоэзді қуатандыратың заттар. Препараттары - әсер ету механизмі, қолдану зәр айдағыштар Варикозное расширение вен. Профилактика и методики оздоровления
Варикозное расширение вен. Профилактика и методики оздоровления Тест 8-ми влечений Сонди и его модификации
Тест 8-ми влечений Сонди и его модификации Темперамент. Типы темперамента и их психологическая характеристика
Темперамент. Типы темперамента и их психологическая характеристика Девиантное поведение. Коррекция
Девиантное поведение. Коррекция Грибковые заболевания кожи. Микозы волосистой части головы
Грибковые заболевания кожи. Микозы волосистой части головы Тіс-жақ жүйесінің дамуы және деформациясы қалыптасуына тамақтанудың рөлі
Тіс-жақ жүйесінің дамуы және деформациясы қалыптасуына тамақтанудың рөлі Школа здоровья для беременных. Занятие 6
Школа здоровья для беременных. Занятие 6 Женские половые гормоны
Женские половые гормоны Развитие беременности и изменения, происходящие в организме женщины во время беременности. Лекция № 1
Развитие беременности и изменения, происходящие в организме женщины во время беременности. Лекция № 1 Возрастная периодизация психического развития человека
Возрастная периодизация психического развития человека ЭКГ при инфаркте миокарда
ЭКГ при инфаркте миокарда Витамины В1, В2 , В6
Витамины В1, В2 , В6 Григорий Антонович Захарьин
Григорий Антонович Захарьин Первичный билиарный цирроз
Первичный билиарный цирроз