- Энзимопатии

Содержание
- 2. Активная регуляция- регуляция на геномном уровне ( лактозный оперон); Регуляция конститутивными и индуцибельными ферментами. Синтез первых
- 3. Гормональная регуляция активности ферментов Осуществляется при: -функционировании АЦ комплекса, - с помощью вторичных посредников- цАМФ, цГМФ,
- 4. Аденилатциклазный механизм
- 5. Некоторые клеточные ответы на действие гормонов, опосредуемое циклическим АМФ
- 6. ИФ3 обеспечивает повышение уровня кальция
- 8. Сигнальные роли ДАГ Если ИФ3 повышает концентрацию ионов Са2+ в цитозоле, другой продукт расщепления фосфатидилинозитол -4,5-
- 9. Роль ПКС в регуляции транскрипции
- 10. Известны два внутриклеточных пути, через которые активированная С-киназа усиливает транскрипцию генов. В одном, С-киназа активирует протеинкиназный
- 11. Взаимосвязь цАМФ и ИФ3 зависимых путей
- 12. Аллостерическая регуляция Allos- иной. Аллостерическая регуляция имеет два определения. Регуляция с помощью другого вещества, не являющегося
- 13. Аллостерические ферменты Это олигомеры с четным числом протомеров Имеют ось симметрии Обычно имеют два центра связывания
- 14. Изоферменты
- 15. Это генетически детерминированные множественные формы ферментов. Тетрамерный фермент, состоящий из двух кодируемых генетически субъединиц А и
- 16. Метаболизм ферментов Превращение ферментов сходно с обменом других белков. Определенный уровень ферментов в тканях поддерживается их
- 18. Имея высокую молекулярную массу, большая часть ферментов не экскретируется с мочой. Ферменты экскретируются с желчью (
- 19. Активность фермента . Активность - это изменение количества субстрата под влиянием фермента в единицу времени. Под
- 20. В практике лабораторий широко пользуются понятием удельная активность. Для этого число cтандартных единиц пересчитывают на какую-либо
- 21. Пример для вычисления активности фермента Исходные данные:25.0 x 10-3 моль л--1 пептида-субстрата, объем реакционной смеси 2.5
- 22. Изменение активности ферментов у больного ИМ в разные сроки заболевания
- 23. Номенклатура и классификация ферментов
- 24. Оксидоредуктазы Оксидоредуктазы (1.0.0.0.) 1.1.0.0. Действуют на СН-ОН группы доноров 1.1.1.0. НАД+ или НАДФ+ в качестве акцепторов
- 25. Трансферазы Трансферазы (2.0.0.0.)2.1.0.0.Переносят одноуглеродные группы 2.1.1.0. Метилтрансферазы 2.1.1.1. Никотинамид метилтрансфераза 2.1.1.45. Тимидилат синтаза 2.3.0.0. Ацилтрансферазы 2.3.1.6.
- 26. Гидролазы (3.0.0.0.)3.1.0.0.Действуют на эфирные связи 3.1.1.0.Гидролазы эфиров карбоновых кислот 3.1.1.17. Ацетилхолинэстераза 3.2.1.0. Гликозидгидролазы 3.2.1.1. a-амилаза 3.2.1.2.
- 27. Лиазы(4.0.0.0)4.1.0.0.Углерод-углерод лиазы 4.1.1.0.Карбокси лиазы 4.1.1.1. Пируватдекарбоксилаза 4.2.0.0. Углерод-кислород-лиазы 4.2.1.0. Гидролиазы 4.2.1.11. Енолаза 4.2.1.12. Фосфоглюконатдегидраза
- 28. Лигазы (6.0.0.0)6.1.0.0. Образуют С-О связи 6.1.1.0.Образуют молекулы аминоацил-тРНК и родственные им соединения. 6.1.1.1. Тирозил-тРНК синтаза 6.5.0.0.
- 29. Каждый фермент получает специфический кодовый номер-шифр фермента, отражающий его положение в классификации: первая цифра характеризует класс
- 31. Скачать презентацию

Активная регуляция- регуляция на геномном уровне ( лактозный оперон);
Регуляция конститутивными и
Активная регуляция- регуляция на геномном уровне ( лактозный оперон);
Регуляция конститутивными и

Гормональная регуляция активности ферментов
Осуществляется при:
-функционировании АЦ комплекса,
- с помощью вторичных
Гормональная регуляция активности ферментов
Осуществляется при:
-функционировании АЦ комплекса,
- с помощью вторичных
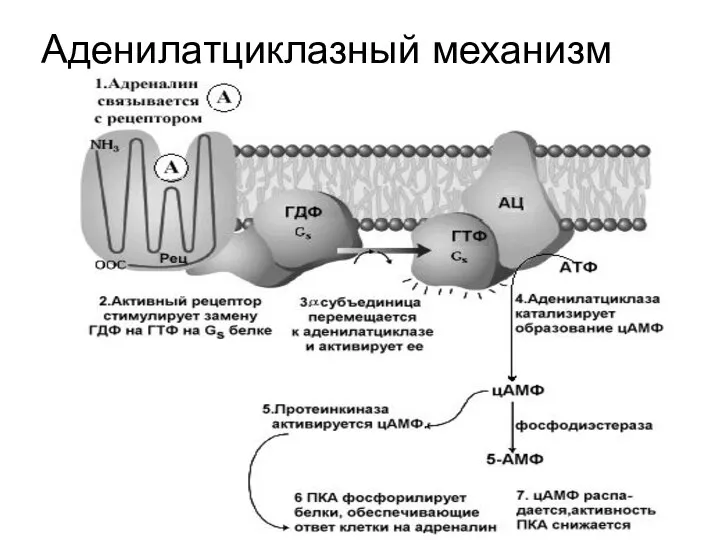
Аденилатциклазный механизм
Аденилатциклазный механизм
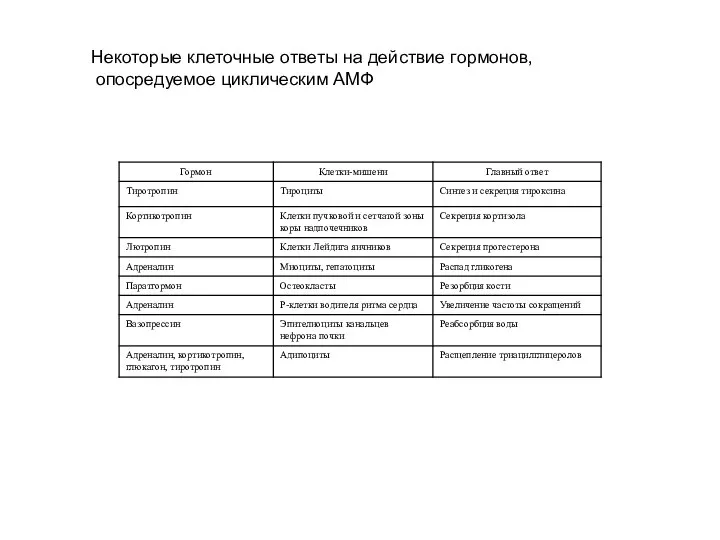
Некоторые клеточные ответы на действие гормонов,
опосредуемое циклическим АМФ
Некоторые клеточные ответы на действие гормонов,
опосредуемое циклическим АМФ
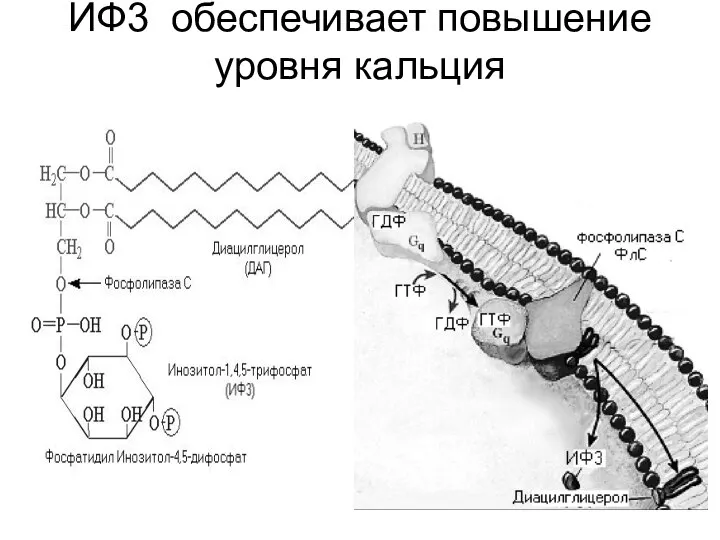
ИФ3 обеспечивает повышение уровня кальция
ИФ3 обеспечивает повышение уровня кальция


Сигнальные роли ДАГ
Если ИФ3 повышает концентрацию ионов Са2+ в цитозоле,
Сигнальные роли ДАГ
Если ИФ3 повышает концентрацию ионов Са2+ в цитозоле,
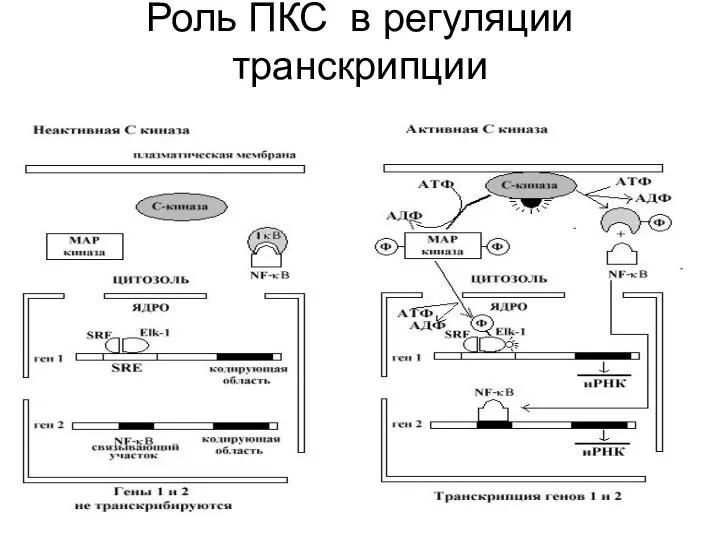
Роль ПКС в регуляции транскрипции
Роль ПКС в регуляции транскрипции

Известны два внутриклеточных пути, через которые активированная С-киназа усиливает транскрипцию генов.
Известны два внутриклеточных пути, через которые активированная С-киназа усиливает транскрипцию генов.
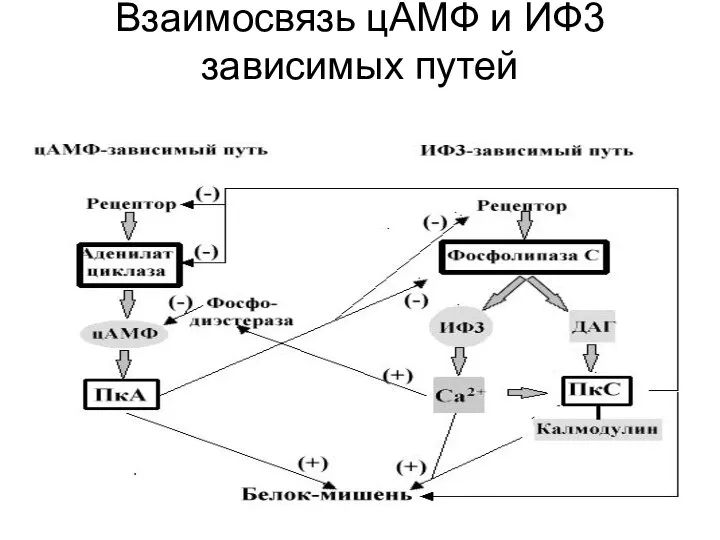
Взаимосвязь цАМФ и ИФ3 зависимых путей
Взаимосвязь цАМФ и ИФ3 зависимых путей

Аллостерическая регуляция
Allos- иной. Аллостерическая регуляция имеет два определения.
Регуляция с помощью другого
Аллостерическая регуляция
Allos- иной. Аллостерическая регуляция имеет два определения.
Регуляция с помощью другого

Аллостерические ферменты
Это олигомеры с четным числом протомеров
Имеют ось симметрии
Обычно имеют два
Аллостерические ферменты
Это олигомеры с четным числом протомеров
Имеют ось симметрии
Обычно имеют два
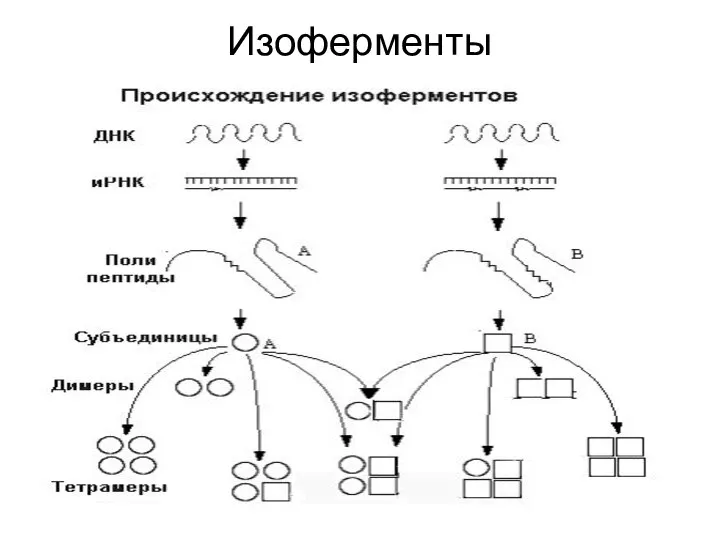
Изоферменты
Изоферменты

Это генетически детерминированные множественные формы ферментов.
Тетрамерный фермент, состоящий из двух кодируемых
Это генетически детерминированные множественные формы ферментов.
Тетрамерный фермент, состоящий из двух кодируемых

Метаболизм ферментов
Превращение ферментов сходно с обменом других белков. Определенный уровень ферментов
Метаболизм ферментов
Превращение ферментов сходно с обменом других белков. Определенный уровень ферментов
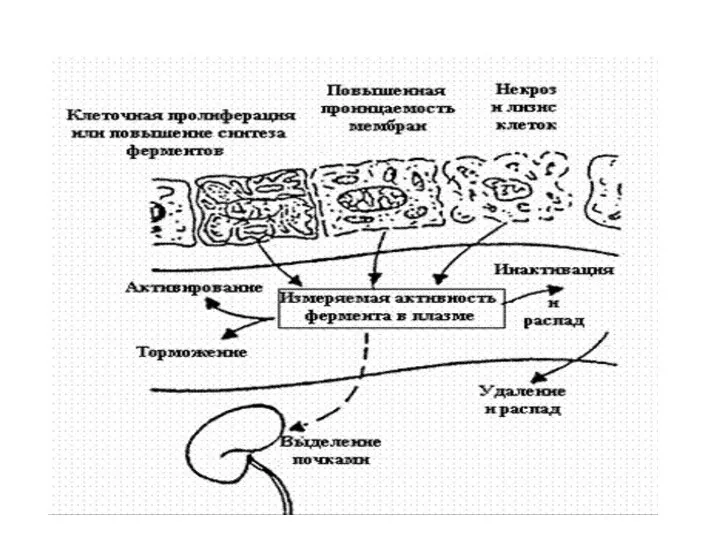

Имея высокую молекулярную массу, большая часть ферментов не экскретируется с мочой.
Имея высокую молекулярную массу, большая часть ферментов не экскретируется с мочой.

Активность фермента
. Активность - это изменение количества субстрата под влиянием фермента
Активность фермента
. Активность - это изменение количества субстрата под влиянием фермента

В практике лабораторий широко пользуются понятием удельная активность. Для этого число
В практике лабораторий широко пользуются понятием удельная активность. Для этого число

Пример для вычисления активности фермента
Исходные данные:25.0 x 10-3 моль л--1 пептида-субстрата,
объем
Пример для вычисления активности фермента
Исходные данные:25.0 x 10-3 моль л--1 пептида-субстрата, объем
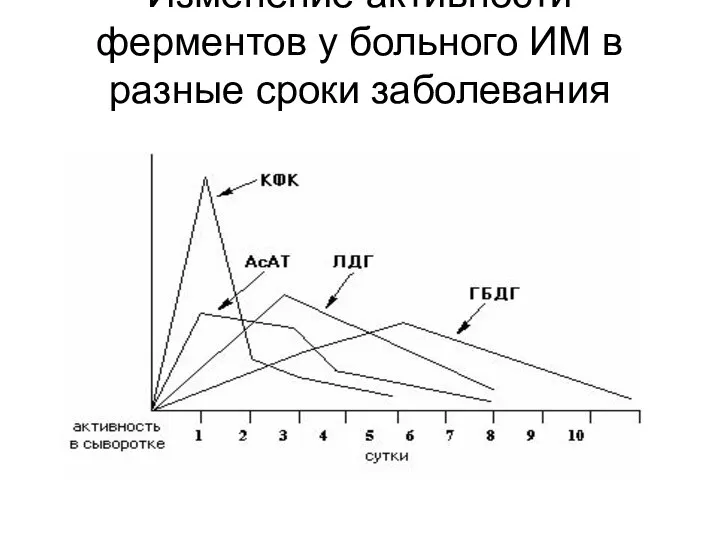
Изменение активности ферментов у больного ИМ в разные сроки заболевания
Изменение активности ферментов у больного ИМ в разные сроки заболевания
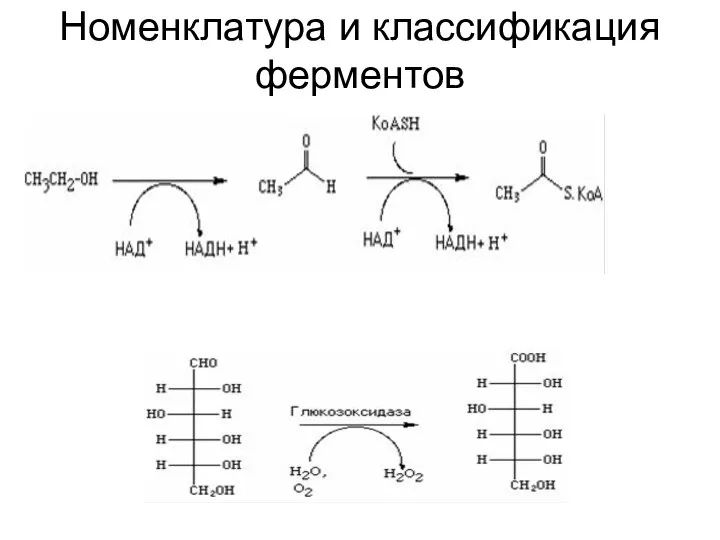
Номенклатура и классификация ферментов
Номенклатура и классификация ферментов

Оксидоредуктазы
Оксидоредуктазы (1.0.0.0.)
1.1.0.0. Действуют на СН-ОН группы доноров
1.1.1.0. НАД+ или НАДФ+
Оксидоредуктазы
Оксидоредуктазы (1.0.0.0.)
1.1.0.0. Действуют на СН-ОН группы доноров
1.1.1.0. НАД+ или НАДФ+

Трансферазы
Трансферазы (2.0.0.0.)2.1.0.0.Переносят одноуглеродные группы 2.1.1.0. Метилтрансферазы 2.1.1.1. Никотинамид метилтрансфераза 2.1.1.45. Тимидилат
Трансферазы
Трансферазы (2.0.0.0.)2.1.0.0.Переносят одноуглеродные группы 2.1.1.0. Метилтрансферазы 2.1.1.1. Никотинамид метилтрансфераза 2.1.1.45. Тимидилат

Гидролазы (3.0.0.0.)3.1.0.0.Действуют на эфирные связи 3.1.1.0.Гидролазы эфиров карбоновых кислот 3.1.1.17. Ацетилхолинэстераза
Гидролазы (3.0.0.0.)3.1.0.0.Действуют на эфирные связи 3.1.1.0.Гидролазы эфиров карбоновых кислот 3.1.1.17. Ацетилхолинэстераза
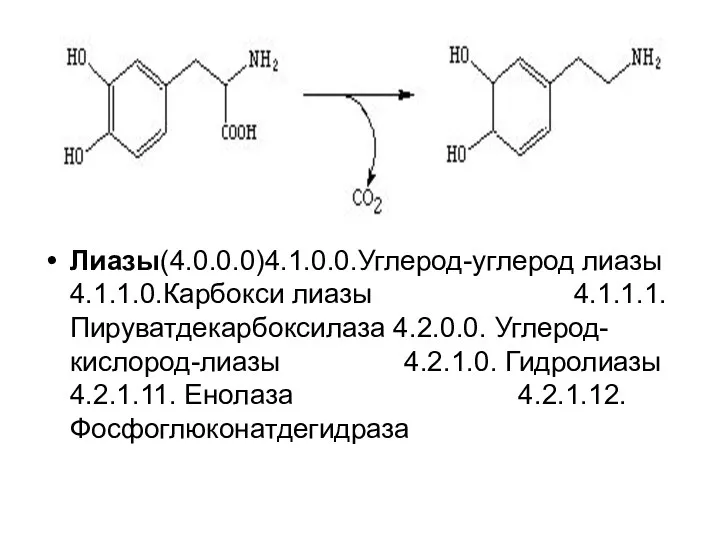
Лиазы(4.0.0.0)4.1.0.0.Углерод-углерод лиазы 4.1.1.0.Карбокси лиазы 4.1.1.1. Пируватдекарбоксилаза 4.2.0.0. Углерод-кислород-лиазы 4.2.1.0. Гидролиазы 4.2.1.11.
Лиазы(4.0.0.0)4.1.0.0.Углерод-углерод лиазы 4.1.1.0.Карбокси лиазы 4.1.1.1. Пируватдекарбоксилаза 4.2.0.0. Углерод-кислород-лиазы 4.2.1.0. Гидролиазы 4.2.1.11.

Лигазы (6.0.0.0)6.1.0.0. Образуют С-О связи 6.1.1.0.Образуют молекулы аминоацил-тРНК и родственные им
Лигазы (6.0.0.0)6.1.0.0. Образуют С-О связи 6.1.1.0.Образуют молекулы аминоацил-тРНК и родственные им

Каждый фермент получает специфический кодовый номер-шифр фермента, отражающий его положение
Каждый фермент получает специфический кодовый номер-шифр фермента, отражающий его положение
 Turystyka kajakowa w Polsce
Turystyka kajakowa w Polsce Web-сайт сети магазинов для видеоигр «GameTime»
Web-сайт сети магазинов для видеоигр «GameTime» «НСОТ – Новая система оплаты труда работников бюджетной сферы» Косова Марина Вячеславовна
«НСОТ – Новая система оплаты труда работников бюджетной сферы» Косова Марина Вячеславовна  Фестиваль детских объединений ГДОО "Росток"
Фестиваль детских объединений ГДОО "Росток" Етнокультурна мозаїка України
Етнокультурна мозаїка України Украинская культура
Украинская культура  Физические лица. Правосубъектность граждан. Тема 3
Физические лица. Правосубъектность граждан. Тема 3 Презентация на тему "Рахит" - скачать презентации по Медицине
Презентация на тему "Рахит" - скачать презентации по Медицине Raspberri Pi, Python и Git. Схема команд
Raspberri Pi, Python и Git. Схема команд Презентация: «Факторы влияющие на принятия управленческих решений» Выполнил: магистр. МС – 11м Нарг
Презентация: «Факторы влияющие на принятия управленческих решений» Выполнил: магистр. МС – 11м Нарг Основы военной связи. Общая тактика
Основы военной связи. Общая тактика Управление карьерой
Управление карьерой Товар Гиффена
Товар Гиффена Основи побудови автоматизованих систем управління військами
Основи побудови автоматизованих систем управління військами Ведение клиентской базы
Ведение клиентской базы Iqtisodiyotning xalqaro integratsiyasida
Iqtisodiyotning xalqaro integratsiyasida Политические роли личности
Политические роли личности УМК «Школа России» Переместительный закон сложения 1 класс
УМК «Школа России» Переместительный закон сложения 1 класс Управление световым оформлением зданий
Управление световым оформлением зданий CAD/CAM/CAE системы. Виды и этапы программирования. Виды моделирования. Уровни CAM систем
CAD/CAM/CAE системы. Виды и этапы программирования. Виды моделирования. Уровни CAM систем Презентация на тему "Система высшего образования новые вызовы и тренды" - скачать презентации по Педагогике
Презентация на тему "Система высшего образования новые вызовы и тренды" - скачать презентации по Педагогике Центробежные компрессоры. Проточный тракт
Центробежные компрессоры. Проточный тракт Цифровой логический уровень. Транзистор
Цифровой логический уровень. Транзистор Презентация на тему "ВОЗРАСНЫЕ ОСОБЕННОСТИ РАЗВИТИЯ ЛИЧНОСТИ" - скачать презентации по Медицине
Презентация на тему "ВОЗРАСНЫЕ ОСОБЕННОСТИ РАЗВИТИЯ ЛИЧНОСТИ" - скачать презентации по Медицине реляционная алгебра SQL
реляционная алгебра SQL Мир верх ногами
Мир верх ногами Программирование на языке Паскаль
Программирование на языке Паскаль Оформление интерьеров Центров образования цифрового и гуманитарного профилей «Точка роста»
Оформление интерьеров Центров образования цифрового и гуманитарного профилей «Точка роста»