- Потенциал действия аксона, его фазы и ионная природа

Содержание
- 2. Изменение мембранного потенциала клетки при действии электрического тока различной силы Действие допорогового стимула Действие порогового стимула
- 3. ПОТЕНЦИАЛ ДЕЙСТВИЯ В КЛЕТКАХ РАЗНЫХ ТКАНЕЙ а б в а – гигантский аксон кальмара; б –
- 4. ХАРАКТЕРИСТИКИ ПОТЕНЦИАЛА ДЕЙСТВИЯ КЛЕТОК РАЗНЫХ ТКАНЕЙ Длительность потенциала действия: 0,5 - 1 мс (нервные клетки) несколько
- 5. Изменение мембранного потенциала Д – фаза деполяризации, РБ – фаза быстрой реполяризации, РМ – фаза медленной
- 6. ИЗМЕНЕНИЕ ИОННОЙ ПРОНИЦАЕМОСТИ ВО ВРЕМЯ РАЗВИТИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ ПП PK: PNa: PCl=1:0,04:0,45 ПД PK: PNa: PCl=
- 7. Изменение проницаемости мембраны для ионов натрия и калия во время потенциала действия
- 8. ВЛИЯНИЕ СОДЕРЖАНИЯ ИОНОВ НАТРИЯ НА АМПЛИТУДУ ПД
- 10. ИОННЫЙ МЕХАНИЗМ ПД
- 12. РЕГЕНЕРАТИВНЫЕ СВЯЗИ МЕЖДУ ДЕПОЛЯРИЗАЦИЕЙ МЕМБРАНЫ, УВЕЛИЧЕНИЕМ НАТРИЕВОЙ ПРОНИЦАЕМОСТИ И И ВХОДЯЩИМ ТОКОМ ИОНОВ НАТРИЯ
- 13. Воротные механизмы потенциалозависимых ионных каналов
- 14. Модель Na+–канала в мембране
- 15. Изменение состояния натриевых и калиевых каналов мембраны в зависимости от величины мембранного потенциала
- 16. Cостояниe ионных каналов в фазу реполяризации ПД
- 20. Схема работы ворот потенциалозависимых натриевых каналов нервной мембраны а - зависимость процента открытых инактивационных (h) и
- 21. МЕТОД ФИКСАЦИИ ПОТЕНЦИАЛА
- 22. Методика фиксации мембранного потенциала (МП) и регистрации трансмембранных токов (Iм) Ус — усилитель, реагирующий выходным током
- 24. ОБЩИЙ МЕМБРАННЫЙ ТОК Im i Емкостной Общий ионный ток ток
- 25. Ионные токи, зарегистрированные методом фиксации потенциала А - ток, протекающий через мембрану (синяя кривая) при смещении
- 26. А - смещения мембранного потенциала во времени относительно поддерживаемого потенциала. Б - ток через мембрану, регистрируемый
- 27. Избирательное блокирование натриевых и калиевых каналов с помощью тетродотоксина и тетраэтиламмония
- 28. МЕТОД patch-clamp и его конфигурации для измерения токов через одиночные каналы
- 29. Na+-ток через одиночный Na+-канал в мышечной клетке мыши. А - Регистрация методом patch-clamp в конфигурации cell-attached
- 30. А - Na+-токи, зарегистрированные у разных электровозбудимых клеток Б - Na+-токи, зарегистрированные при разных ступеньках относительно
- 31. ФОРМАЛЬНОЕ ОПИСАНИЕ ИОННЫХ ТОКОВ (МОДЕЛЬ ХОДЖКИНА - ХАКСЛИ)
- 32. Ионный ток (Ii) складывается из суммы натриевого (INa) калиевого (IK) и тока утечки (IL): Каждый из
- 33. где: gNa, gK и gL – проводимости для ионов натрия, калия и ионов утечки, соответственно (V-V(Na,K,L))
- 34. УРАВНЕНИЯ ХОДЖКИНА - ХАКСЛИ gNa, gK – максимальные проводимости мембраны при сильной деполяризации
- 35. Величины m, h и n рассчитываются из системы дифференциальных уравнений:
- 36. Величины αm, βm, αn, βn, αh, βh – константы скоростей, зависящие от мембранного потенциала, температуры и
- 37. Решения этих уравнений проще представить в виде экспоненциальных характеристик – постоянных времени изменения m, n и
- 38. Графики зависимости стационарных значений m, n и h (m∞, n∞ и h∞) и постоянных времени τm,τn
- 40. Сравнение рассчитанного потенциала действия (В1) с реальным потенциалом действия, зарегистрированным в гигантском аксоне кальмара (В2). Рассчитанная
- 41. Электрические параметры нервных волокон
- 42. Если С=0,8мкФ/см2, ε = 2, то d=2,2 нм Удельное сопротивление аксоплазмы и саркоплазмы от 30 до
- 43. КАБЕЛЬНАЯ СТРУКТУРА Cm емкость мембраны rm сопротивление мембраны ri – сопротивление аксоплазмы
- 44. Согласно закону Ома ток, текущий по осевому цилиндру: i – ток, текущий по осевому цилиндру, х
- 45. Выразим i: (2)
- 46. Ток через мембрану Исходя из 1 и 2 (3) (4)
- 47. Мембранный ток складывается из двух компонентов: (5)
- 48. Далее из 4 и 5 получаем Умножим обе части на rm (6) (7)
- 49. Заменим (8) Получим
- 50. КАБЕЛЬНОЕ УРАВНЕНИЕ
- 51. Решением этого уравнения является экспоненциальная зависимость: Где τ– постоянная времени, показывающая через сколько времени амплитуда мембранного
- 52. Если продолжительность прямоугольного толчка тока превышает 3τ, V достигает постоянного уровня, в этом случае Уравнение примет
- 53. Решение этого уравнения:
- 54. Сопротивление мембраны (rm) рассчитывается через удельное сопротивление (Rm): где Rm=1-100 кОм·см2, в перехватах Ранвье: 30-40 Ом·см2
- 55. λ– постоянная длины, показывающая на каком расстоянии амплитуда мембранного потенциала падает в e раз. с учетом
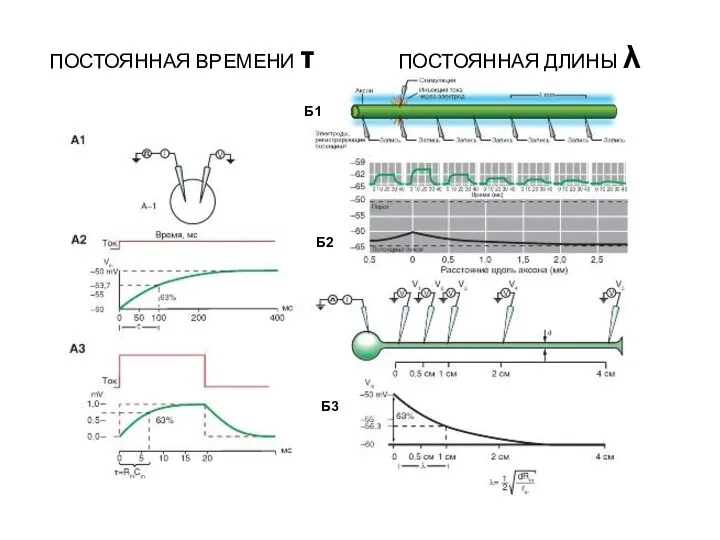
- 56. Б1 Б2 Б3 ПОСТОЯННАЯ ВРЕМЕНИ τ ПОСТОЯННАЯ ДЛИНЫ λ
- 57. Распространение пассивного электротонического потенциала на примере немиелинизированного волокна. Na канал обозначен розовым цветом К канал –
- 58. Типы нервных волокон А - миелиновое волокно, Б - безмиелиновое волокно. 1 - осевой цилиндр, 2
- 59. Распространение потенциала действия на примере немиелинизированного волокна. Na каналы обозначены розовым цветом К каналы – желтым
- 60. Механизм проведения потенциала действия в немиелинизированном волокне.
- 61. Механизм распространения возбуждения по безмиелиновому нервному волокну
- 62. Изменение мембранного потенциала и натриевой и калиевой проводимости мембраны вдоль кабельной структуры во время распространяющегося ПД,
- 63. Миелиновое нервное волокно
- 64. Механизм проведения потенциала действия в миелинизированном волокне.
- 65. Механизм распространения возбуждения по миелиновому нервному волокну
- 67. Сальтаторное проведение
- 69. Скачать презентацию
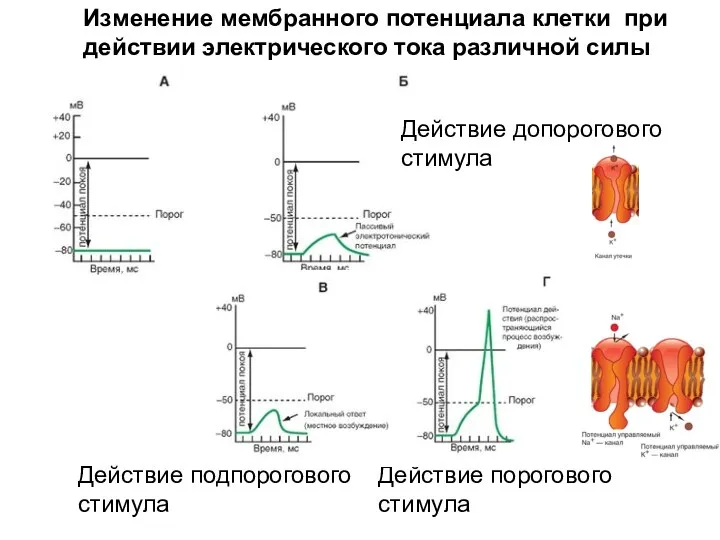
Изменение мембранного потенциала клетки при действии электрического тока различной силы
Действие
Изменение мембранного потенциала клетки при действии электрического тока различной силы
Действие
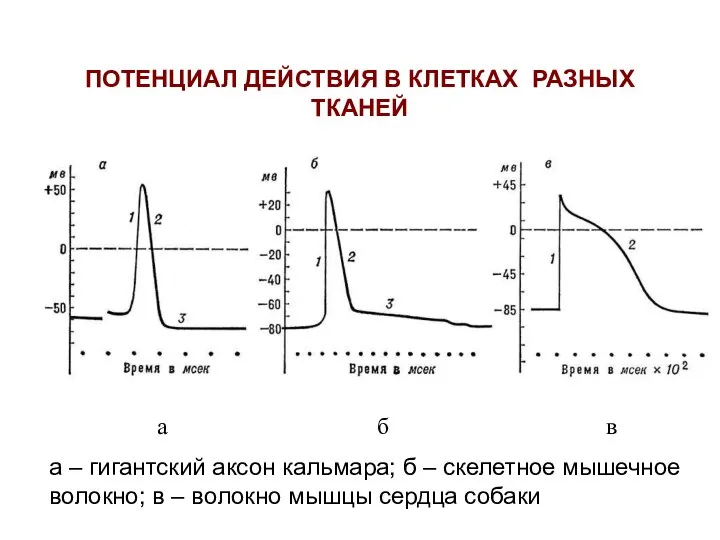
ПОТЕНЦИАЛ ДЕЙСТВИЯ В КЛЕТКАХ РАЗНЫХ ТКАНЕЙ
а б в
а – гигантский
ПОТЕНЦИАЛ ДЕЙСТВИЯ В КЛЕТКАХ РАЗНЫХ ТКАНЕЙ
а б в
а – гигантский

ХАРАКТЕРИСТИКИ ПОТЕНЦИАЛА ДЕЙСТВИЯ КЛЕТОК РАЗНЫХ ТКАНЕЙ
Длительность потенциала действия:
0,5 -
ХАРАКТЕРИСТИКИ ПОТЕНЦИАЛА ДЕЙСТВИЯ КЛЕТОК РАЗНЫХ ТКАНЕЙ
Длительность потенциала действия:
0,5 -
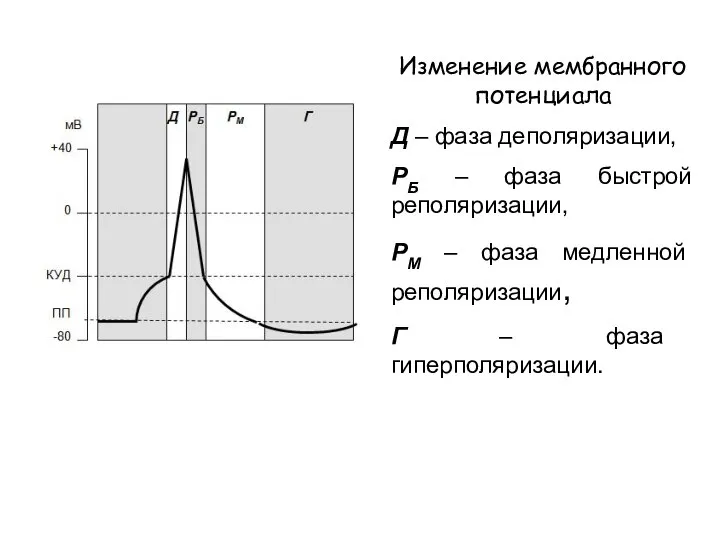
Изменение мембранного потенциала
Д – фаза деполяризации,
РБ – фаза быстрой реполяризации,
Изменение мембранного потенциала
Д – фаза деполяризации,
РБ – фаза быстрой реполяризации,
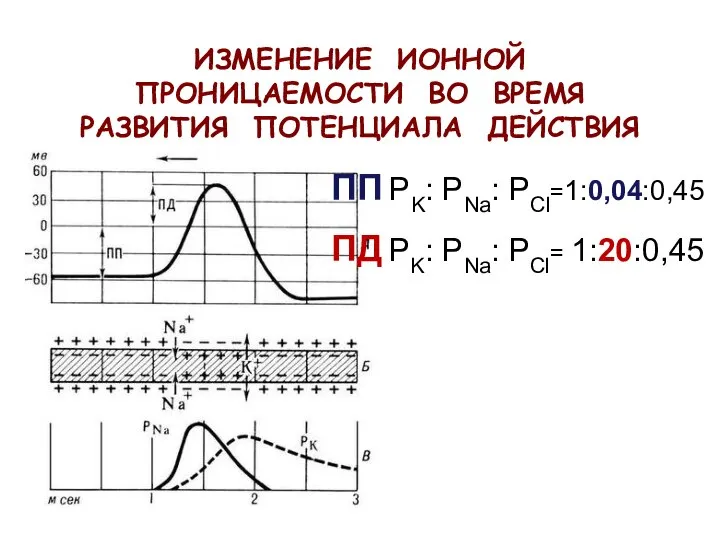
ИЗМЕНЕНИЕ ИОННОЙ ПРОНИЦАЕМОСТИ ВО ВРЕМЯ РАЗВИТИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ
ПП PK: PNa: PCl=1:0,04:0,45
ПД
ИЗМЕНЕНИЕ ИОННОЙ ПРОНИЦАЕМОСТИ ВО ВРЕМЯ РАЗВИТИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ
ПП PK: PNa: PCl=1:0,04:0,45
ПД
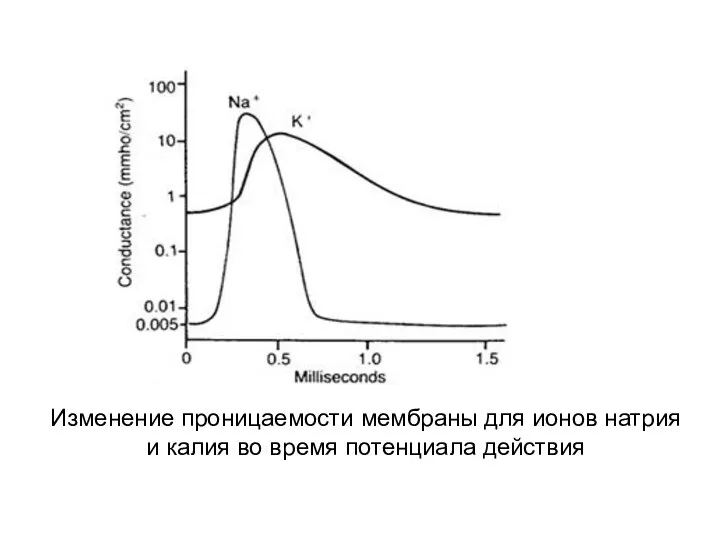
Изменение проницаемости мембраны для ионов натрия и калия во время потенциала
Изменение проницаемости мембраны для ионов натрия и калия во время потенциала
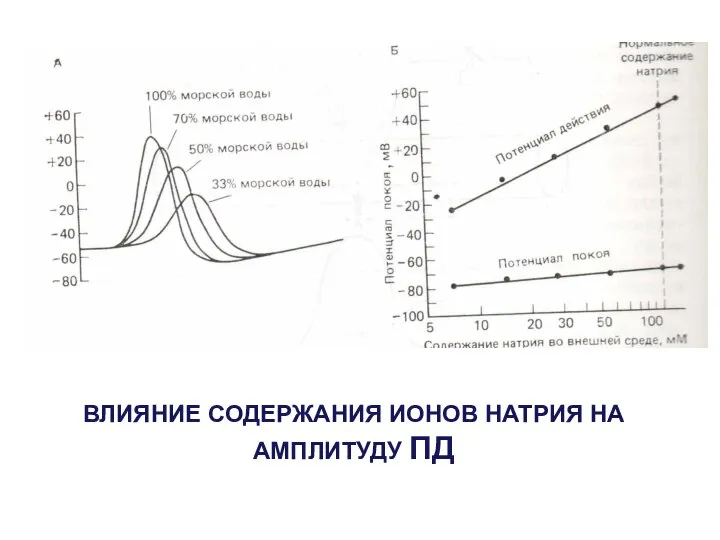
ВЛИЯНИЕ СОДЕРЖАНИЯ ИОНОВ НАТРИЯ НА АМПЛИТУДУ ПД
ВЛИЯНИЕ СОДЕРЖАНИЯ ИОНОВ НАТРИЯ НА АМПЛИТУДУ ПД
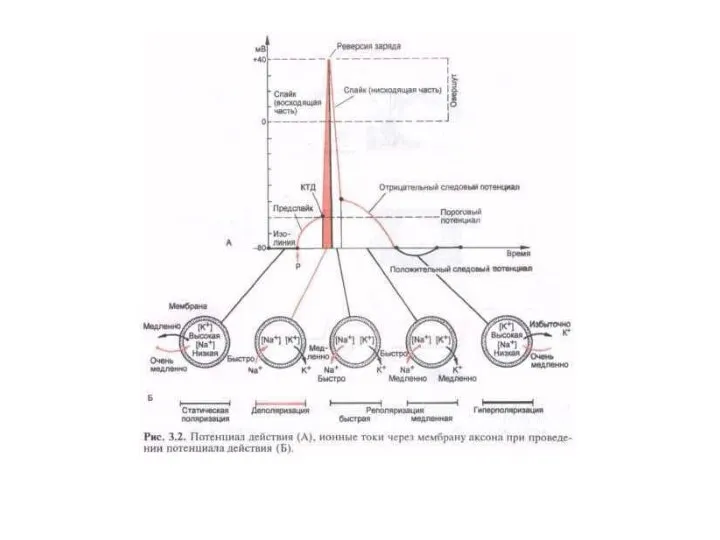
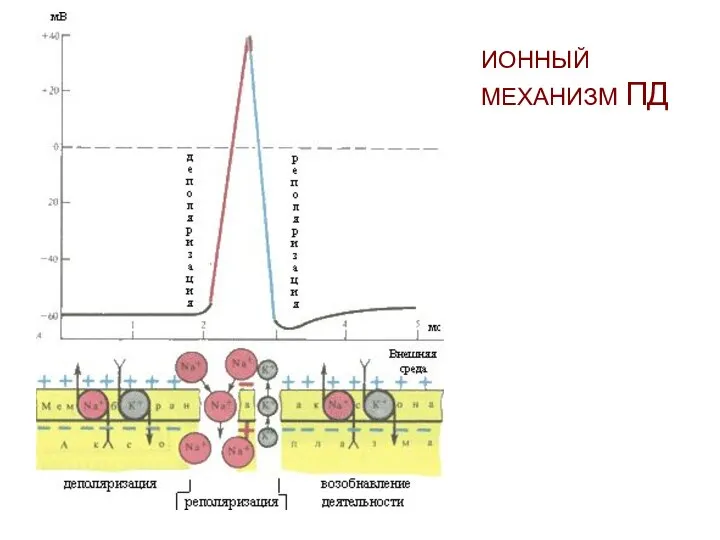
ИОННЫЙ МЕХАНИЗМ ПД
ИОННЫЙ МЕХАНИЗМ ПД
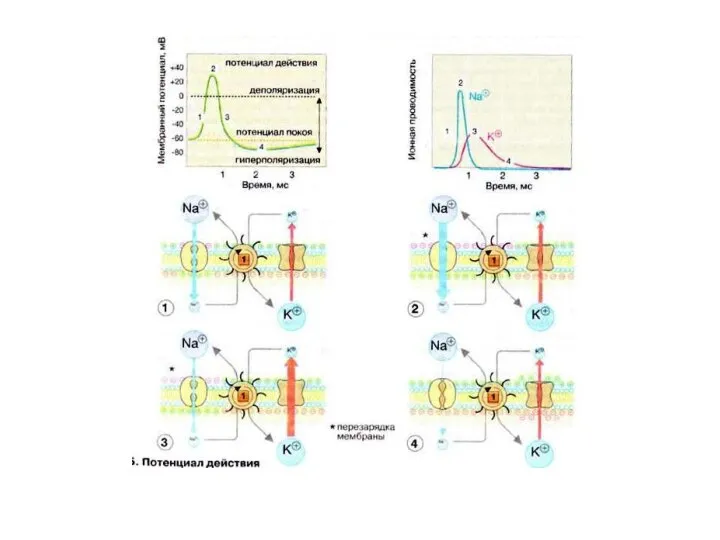

РЕГЕНЕРАТИВНЫЕ СВЯЗИ МЕЖДУ ДЕПОЛЯРИЗАЦИЕЙ МЕМБРАНЫ, УВЕЛИЧЕНИЕМ НАТРИЕВОЙ ПРОНИЦАЕМОСТИ И И ВХОДЯЩИМ
РЕГЕНЕРАТИВНЫЕ СВЯЗИ МЕЖДУ ДЕПОЛЯРИЗАЦИЕЙ МЕМБРАНЫ, УВЕЛИЧЕНИЕМ НАТРИЕВОЙ ПРОНИЦАЕМОСТИ И И ВХОДЯЩИМ

Воротные механизмы потенциалозависимых ионных каналов
Воротные механизмы потенциалозависимых ионных каналов
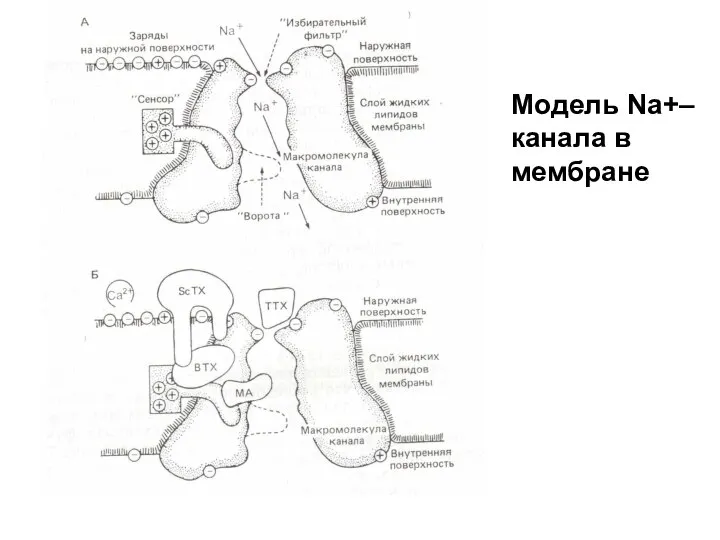
Модель Na+–канала в мембране
Модель Na+–канала в мембране
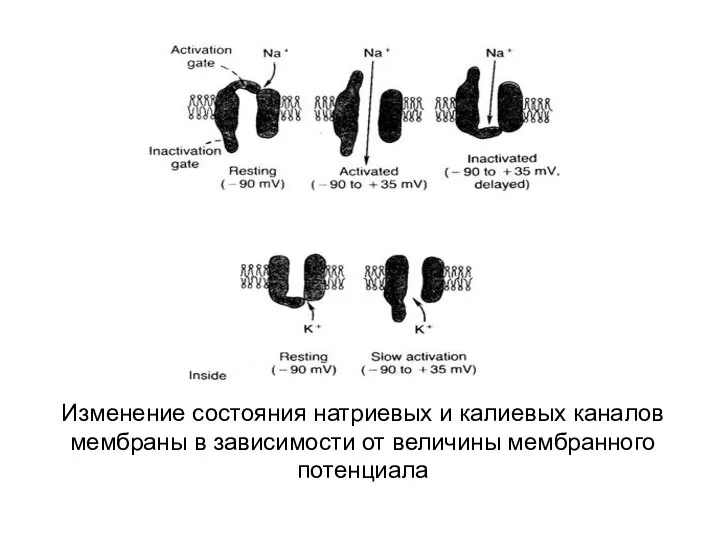
Изменение состояния натриевых и калиевых каналов мембраны в зависимости от величины
Изменение состояния натриевых и калиевых каналов мембраны в зависимости от величины
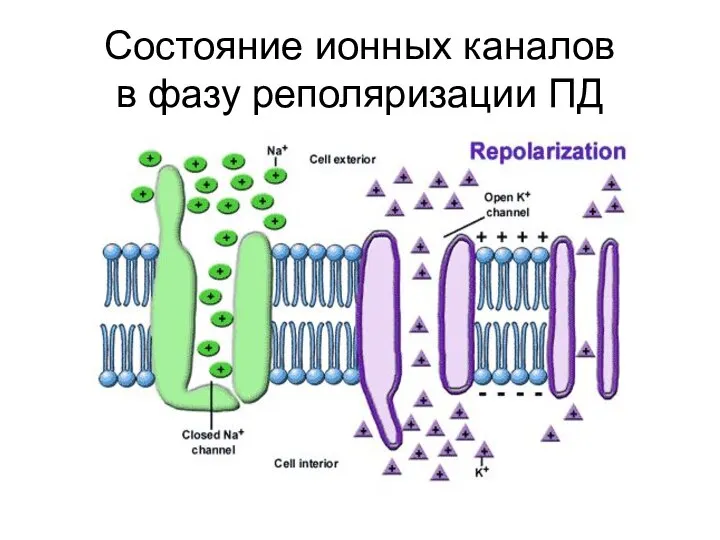
Cостояниe ионных каналов
в фазу реполяризации ПД
Cостояниe ионных каналов
в фазу реполяризации ПД
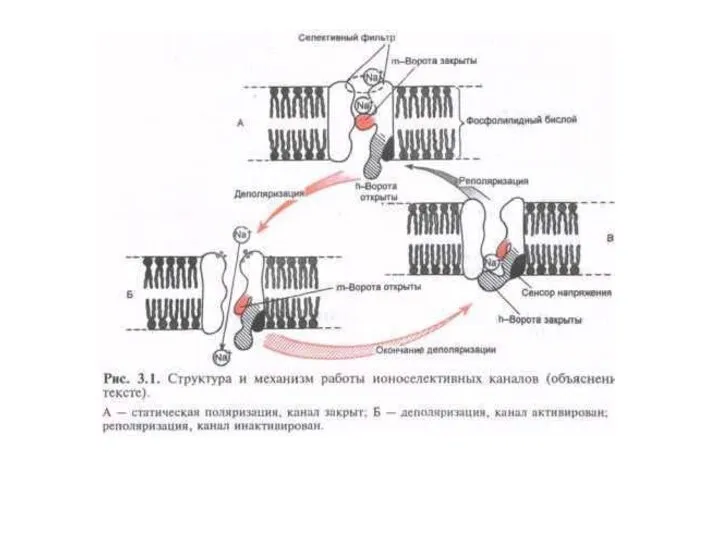
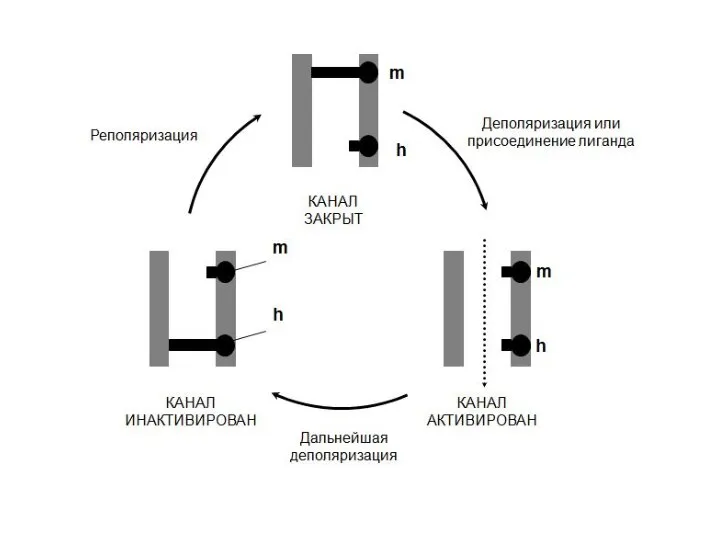
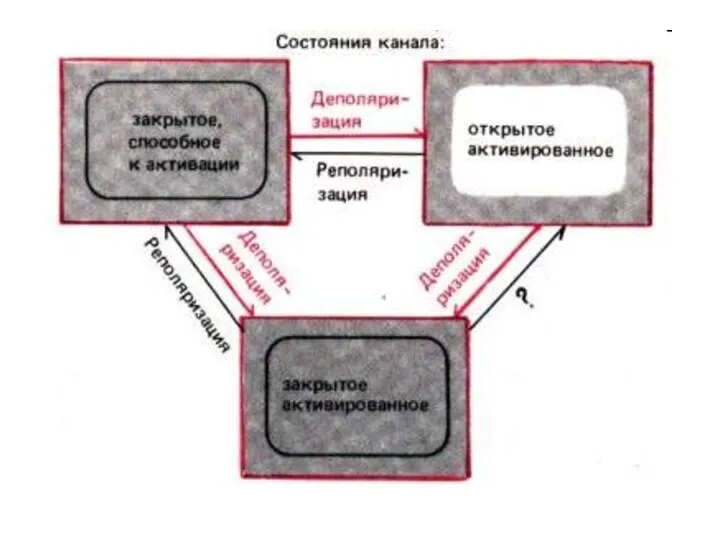
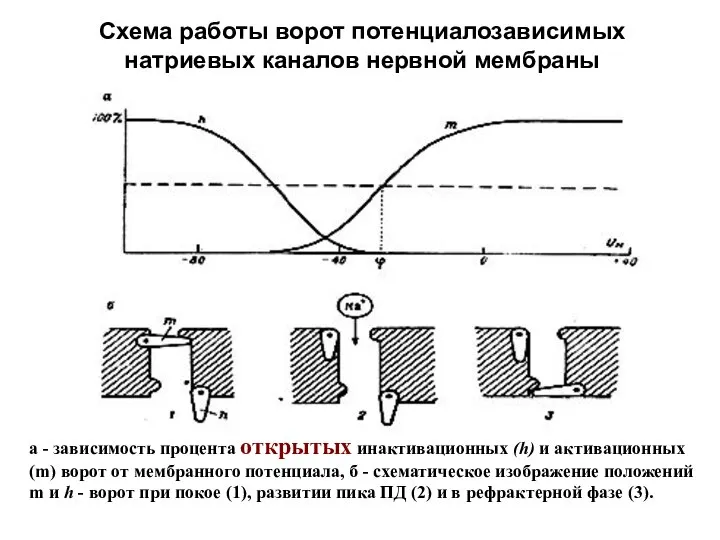
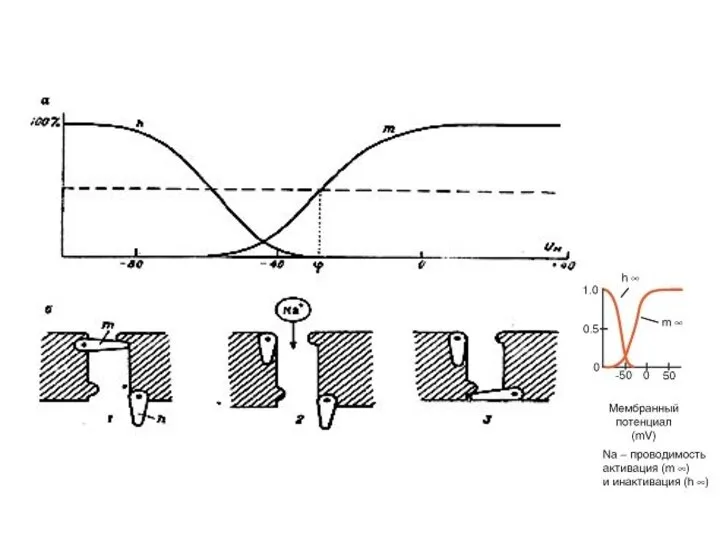
Схема работы ворот потенциалозависимых натриевых каналов нервной мембраны
а - зависимость
Схема работы ворот потенциалозависимых натриевых каналов нервной мембраны
а - зависимость

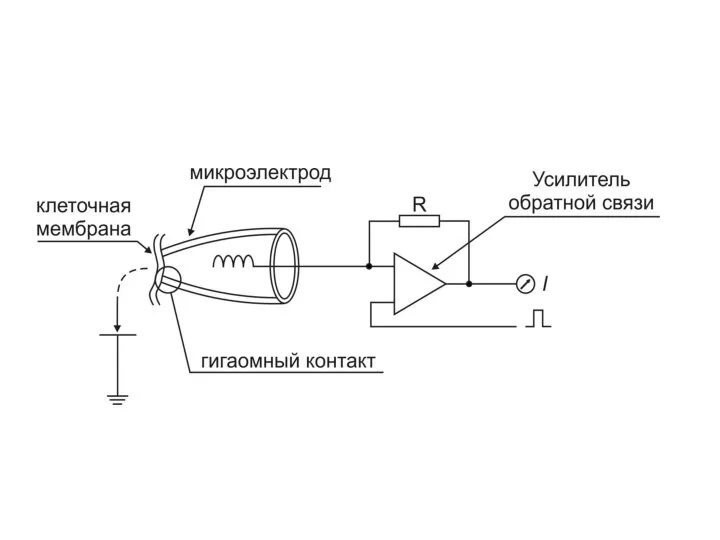
МЕТОД ФИКСАЦИИ ПОТЕНЦИАЛА
МЕТОД ФИКСАЦИИ ПОТЕНЦИАЛА
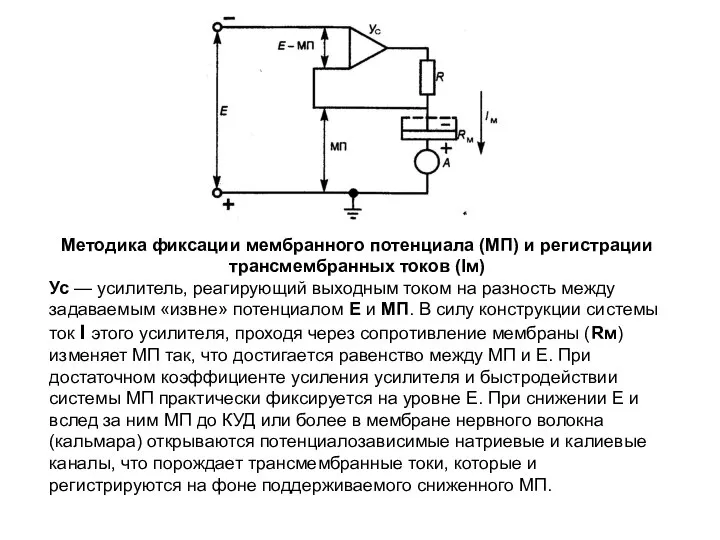
Методика фиксации мембранного потенциала (МП) и регистрации трансмембранных токов (Iм)
Ус —
Методика фиксации мембранного потенциала (МП) и регистрации трансмембранных токов (Iм)
Ус —

ОБЩИЙ МЕМБРАННЫЙ ТОК Im
i
Емкостной Общий ионный ток ток
ОБЩИЙ МЕМБРАННЫЙ ТОК Im
i
Емкостной Общий ионный ток ток
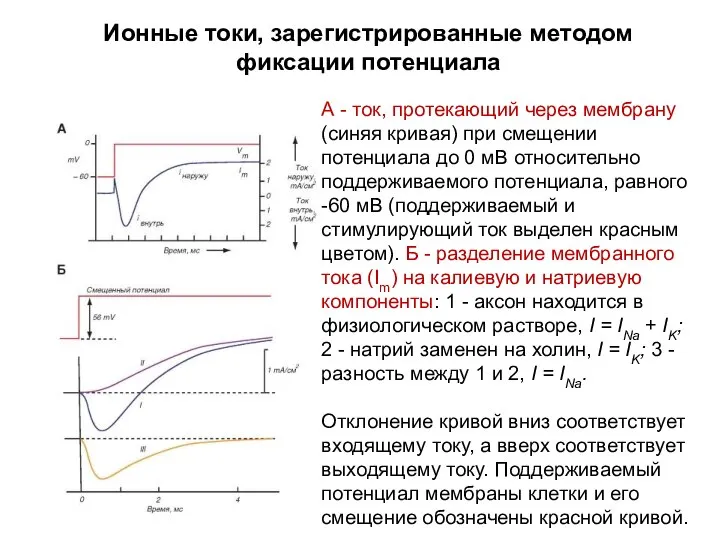
Ионные токи, зарегистрированные методом фиксации потенциала
А - ток, протекающий через мембрану
Ионные токи, зарегистрированные методом фиксации потенциала
А - ток, протекающий через мембрану
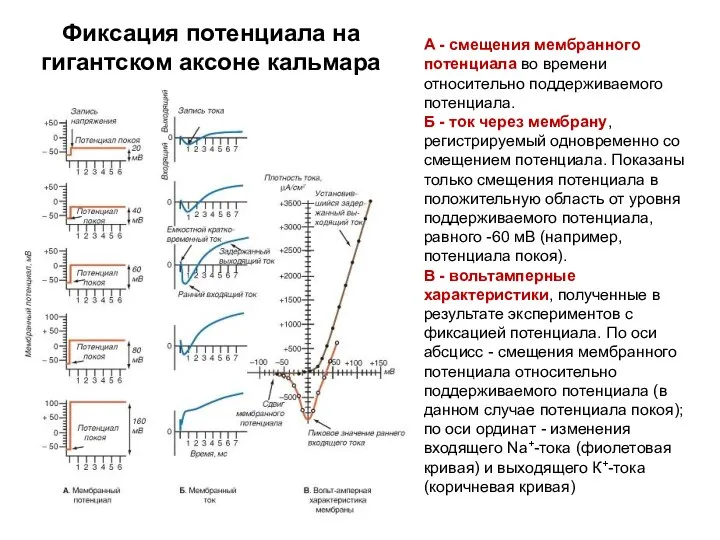
А - смещения мембранного потенциала во времени относительно поддерживаемого потенциала.
Б
А - смещения мембранного потенциала во времени относительно поддерживаемого потенциала.
Б
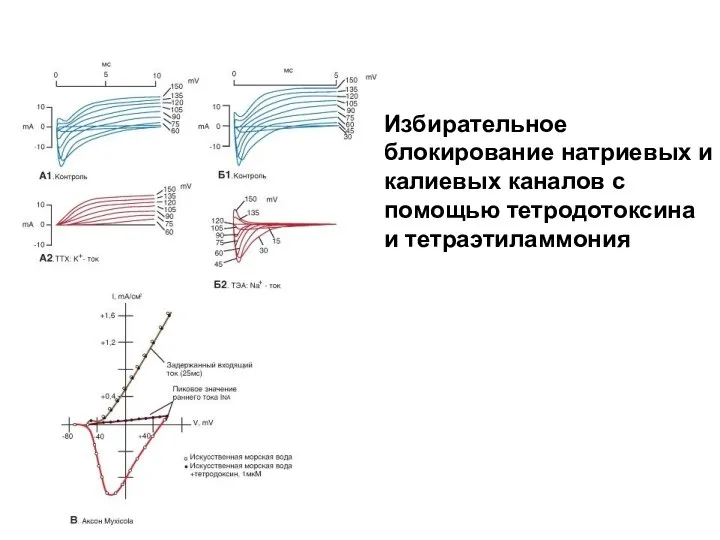
Избирательное блокирование натриевых и калиевых каналов с помощью тетродотоксина и тетраэтиламмония
Избирательное блокирование натриевых и калиевых каналов с помощью тетродотоксина и тетраэтиламмония
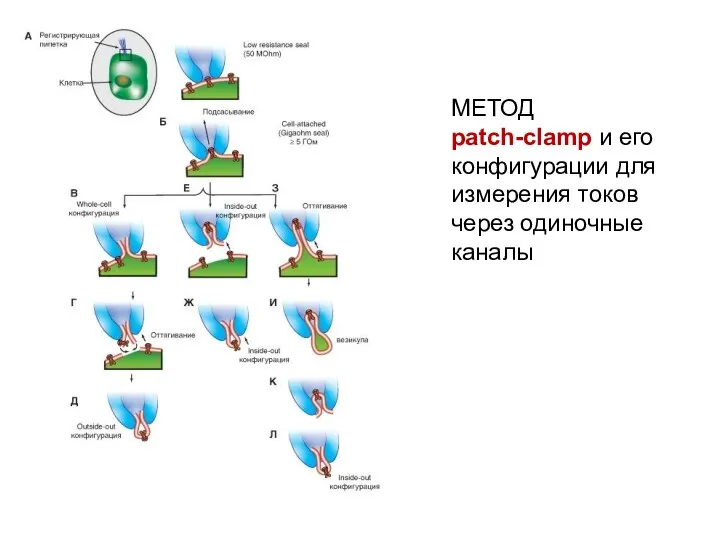
МЕТОД patch-clamp и его конфигурации для измерения токов через одиночные каналы
МЕТОД patch-clamp и его конфигурации для измерения токов через одиночные каналы
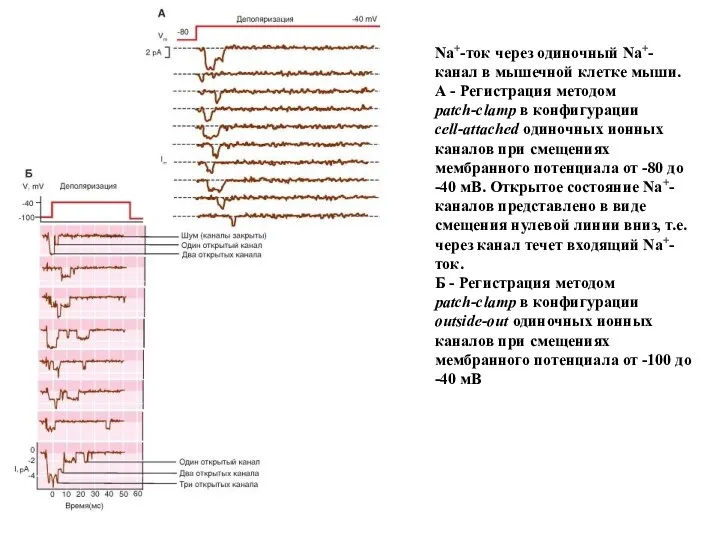
Na+-ток через одиночный Na+-канал в мышечной клетке мыши. А - Регистрация
Na+-ток через одиночный Na+-канал в мышечной клетке мыши. А - Регистрация
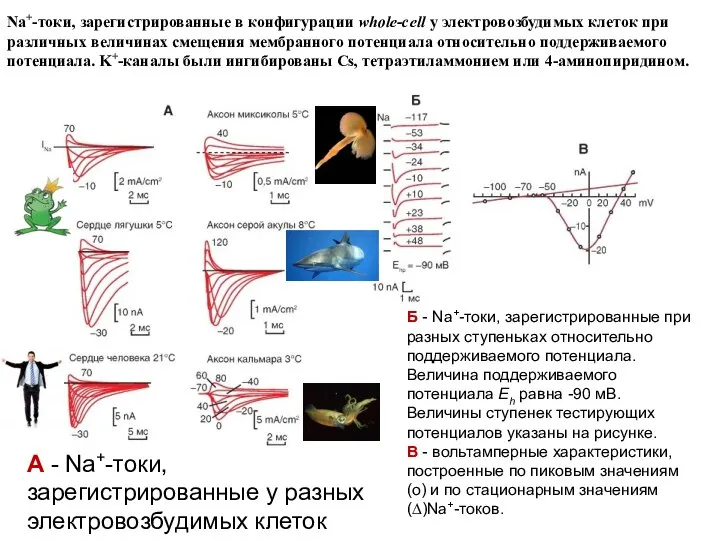
А - Na+-токи, зарегистрированные у разных электровозбудимых клеток
Б - Na+-токи, зарегистрированные
А - Na+-токи, зарегистрированные у разных электровозбудимых клеток
Б - Na+-токи, зарегистрированные

ФОРМАЛЬНОЕ ОПИСАНИЕ ИОННЫХ ТОКОВ (МОДЕЛЬ ХОДЖКИНА - ХАКСЛИ)
ФОРМАЛЬНОЕ ОПИСАНИЕ ИОННЫХ ТОКОВ (МОДЕЛЬ ХОДЖКИНА - ХАКСЛИ)

Ионный ток (Ii) складывается из суммы натриевого (INa) калиевого (IK) и
Ионный ток (Ii) складывается из суммы натриевого (INa) калиевого (IK) и

где:
gNa, gK и gL – проводимости для ионов натрия, калия
где:
gNa, gK и gL – проводимости для ионов натрия, калия

УРАВНЕНИЯ ХОДЖКИНА - ХАКСЛИ
gNa, gK – максимальные проводимости мембраны при сильной
УРАВНЕНИЯ ХОДЖКИНА - ХАКСЛИ
gNa, gK – максимальные проводимости мембраны при сильной

Величины m, h и n рассчитываются из системы дифференциальных уравнений:
Величины m, h и n рассчитываются из системы дифференциальных уравнений:

Величины αm, βm, αn, βn, αh, βh – константы скоростей, зависящие
Величины αm, βm, αn, βn, αh, βh – константы скоростей, зависящие

Решения этих уравнений проще представить в виде экспоненциальных характеристик – постоянных
Решения этих уравнений проще представить в виде экспоненциальных характеристик – постоянных
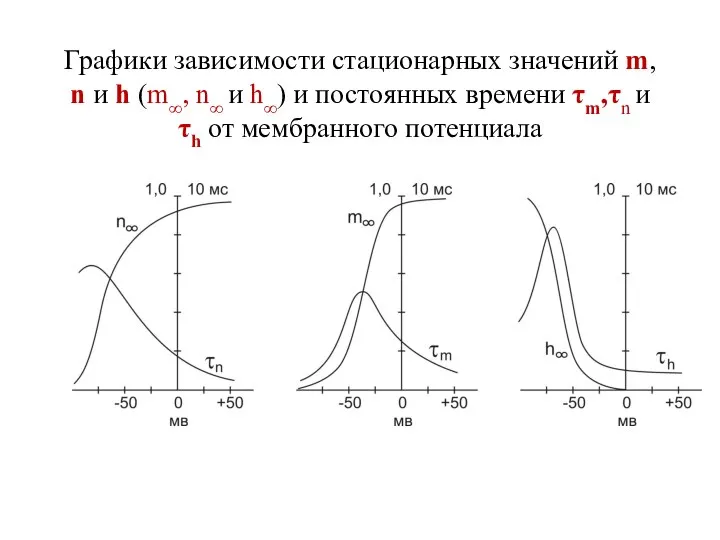
Графики зависимости стационарных значений m, n и h (m∞, n∞ и
Графики зависимости стационарных значений m, n и h (m∞, n∞ и
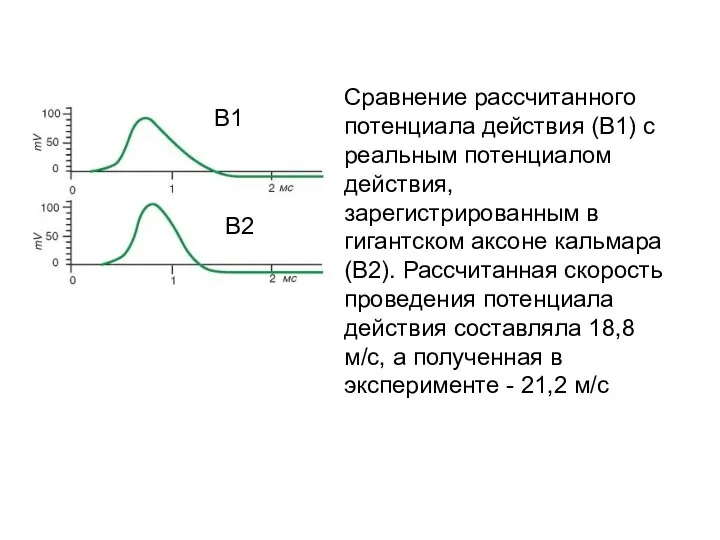
Сравнение рассчитанного потенциала действия (В1) с реальным потенциалом действия, зарегистрированным в
Сравнение рассчитанного потенциала действия (В1) с реальным потенциалом действия, зарегистрированным в

Электрические параметры нервных волокон
Электрические параметры нервных волокон

Если С=0,8мкФ/см2, ε = 2, то d=2,2 нм
Удельное сопротивление аксоплазмы и
Если С=0,8мкФ/см2, ε = 2, то d=2,2 нм
Удельное сопротивление аксоплазмы и
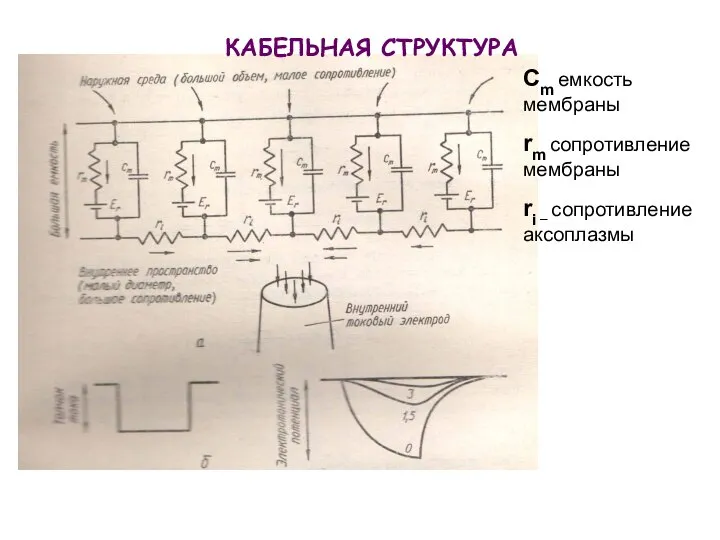
КАБЕЛЬНАЯ СТРУКТУРА
Cm емкость мембраны
rm сопротивление мембраны
ri – сопротивление аксоплазмы
КАБЕЛЬНАЯ СТРУКТУРА
Cm емкость мембраны
rm сопротивление мембраны
ri – сопротивление аксоплазмы

Согласно закону Ома ток, текущий по осевому цилиндру:
i – ток, текущий
Согласно закону Ома ток, текущий по осевому цилиндру:
i – ток, текущий
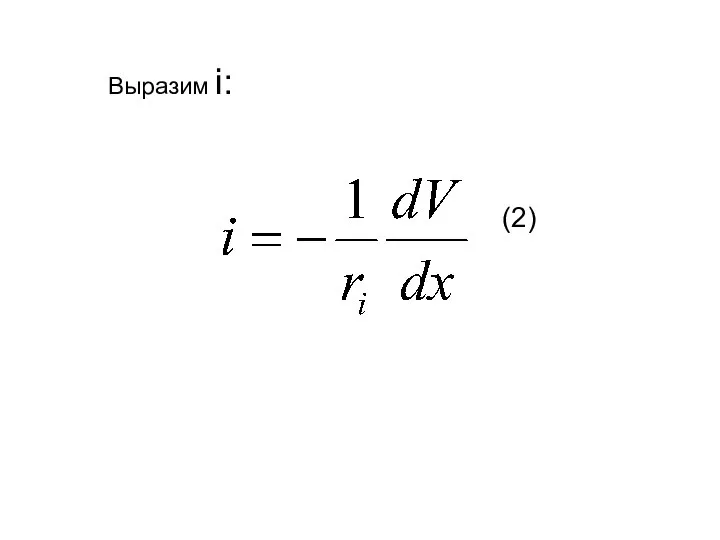
Выразим i:
(2)
Выразим i:
(2)
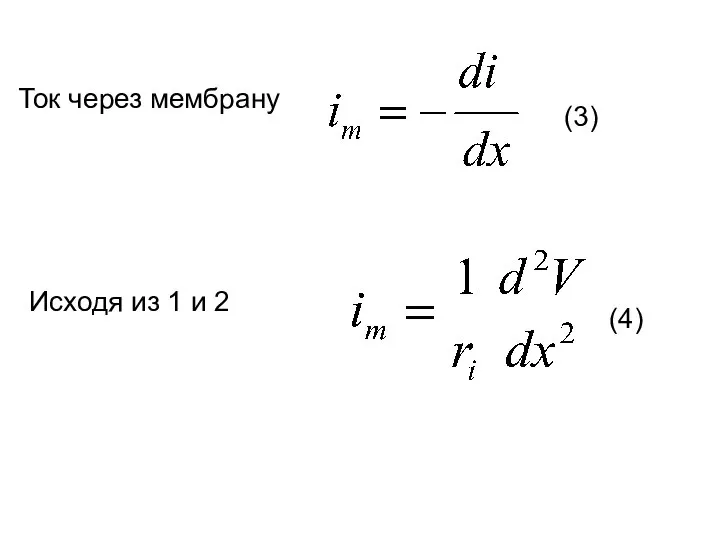
Ток через мембрану
Исходя из 1 и 2
(3)
(4)
Ток через мембрану
Исходя из 1 и 2
(3)
(4)

Мембранный ток складывается из двух компонентов:
(5)
Мембранный ток складывается из двух компонентов:
(5)
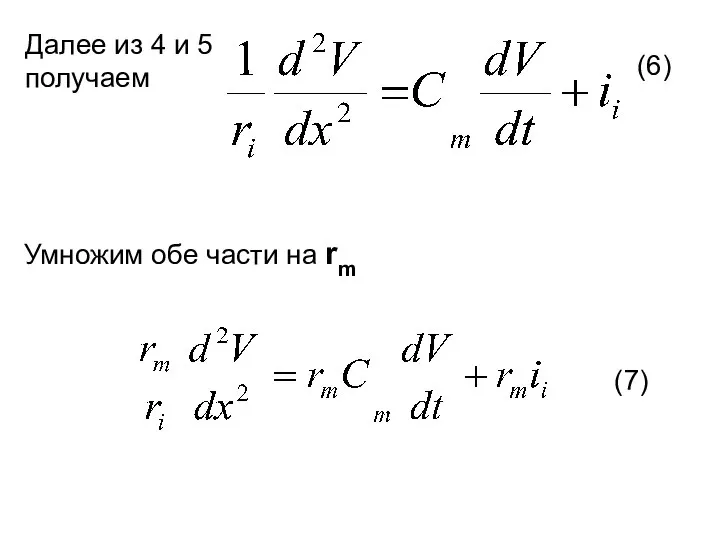
Далее из 4 и 5 получаем
Умножим обе части на rm
(6)
(7)
Далее из 4 и 5 получаем
Умножим обе части на rm
(6)
(7)
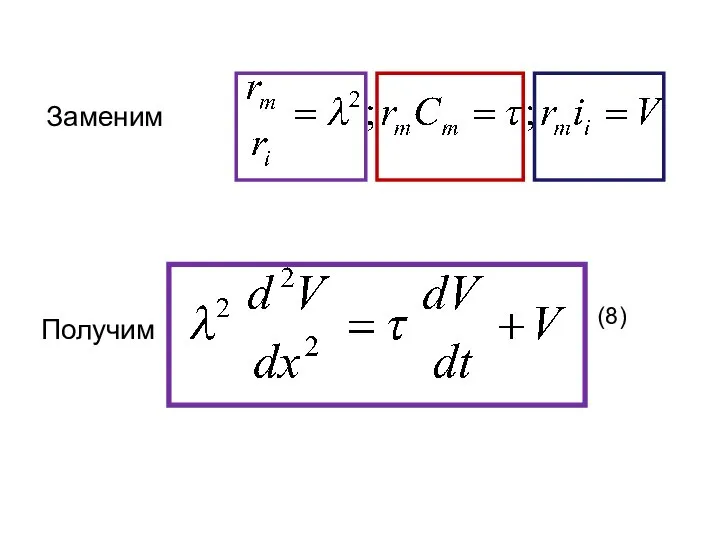
Заменим
(8)
Получим
Заменим
(8)
Получим

КАБЕЛЬНОЕ УРАВНЕНИЕ
КАБЕЛЬНОЕ УРАВНЕНИЕ

Решением этого уравнения является экспоненциальная зависимость:
Где τ– постоянная времени, показывающая через
Решением этого уравнения является экспоненциальная зависимость:
Где τ– постоянная времени, показывающая через

Если продолжительность прямоугольного толчка тока превышает 3τ, V достигает постоянного уровня,
Если продолжительность прямоугольного толчка тока превышает 3τ, V достигает постоянного уровня,

Решение этого уравнения:
Решение этого уравнения:

Сопротивление мембраны (rm) рассчитывается через удельное сопротивление (Rm):
где Rm=1-100 кОм·см2, в
Сопротивление мембраны (rm) рассчитывается через удельное сопротивление (Rm):
где Rm=1-100 кОм·см2, в

λ– постоянная длины, показывающая на каком расстоянии амплитуда мембранного потенциала падает
λ– постоянная длины, показывающая на каком расстоянии амплитуда мембранного потенциала падает
Б1
Б2
Б3
ПОСТОЯННАЯ ВРЕМЕНИ τ ПОСТОЯННАЯ ДЛИНЫ λ
Б1
Б2
Б3
ПОСТОЯННАЯ ВРЕМЕНИ τ ПОСТОЯННАЯ ДЛИНЫ λ
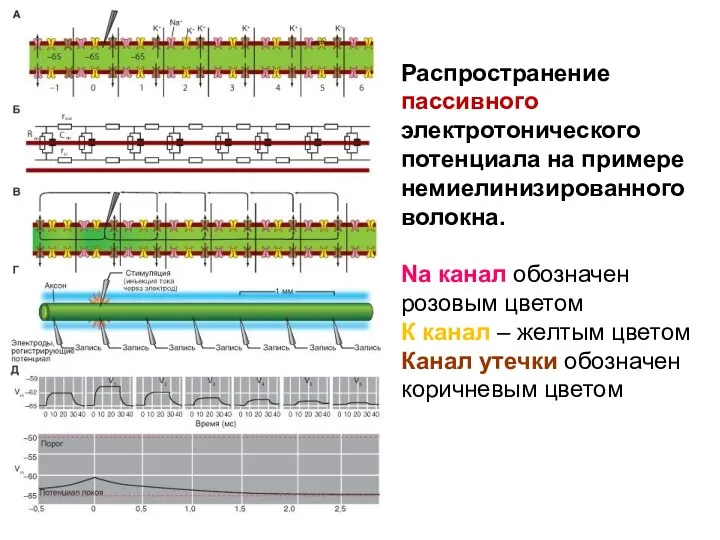
Распространение пассивного электротонического потенциала на примере немиелинизированного волокна.
Na канал обозначен розовым
Распространение пассивного электротонического потенциала на примере немиелинизированного волокна.
Na канал обозначен розовым
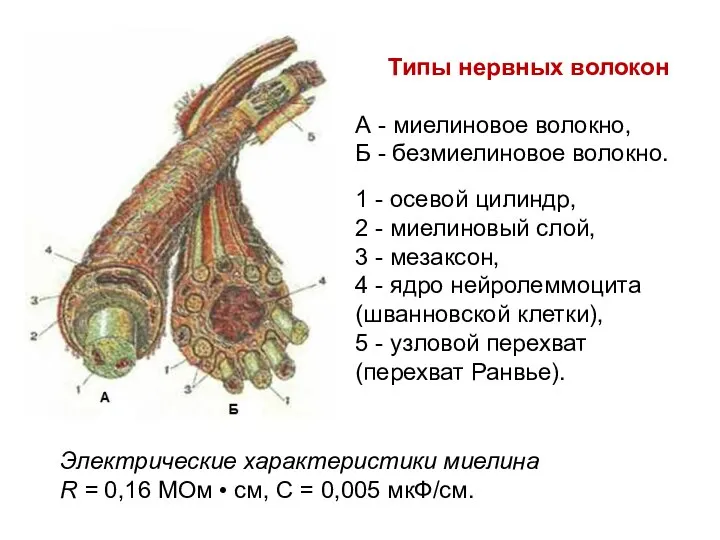
Типы нервных волокон
А - миелиновое волокно,
Б - безмиелиновое волокно.
1 -
Типы нервных волокон
А - миелиновое волокно,
Б - безмиелиновое волокно.
1 -
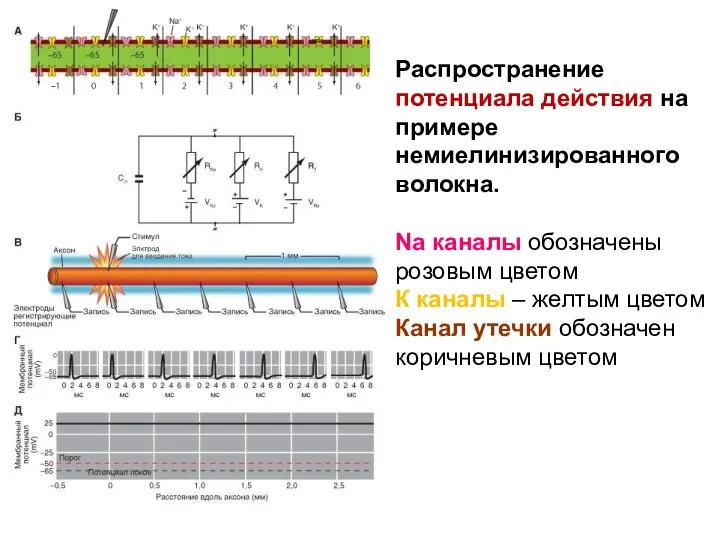
Распространение потенциала действия на примере немиелинизированного волокна.
Na каналы обозначены розовым цветом
К
Распространение потенциала действия на примере немиелинизированного волокна.
Na каналы обозначены розовым цветом
К
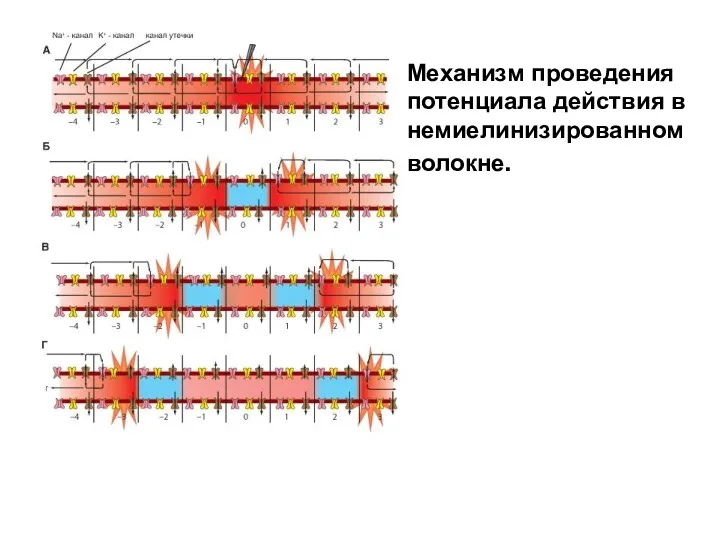
Механизм проведения потенциала действия в немиелинизированном волокне.
Механизм проведения потенциала действия в немиелинизированном волокне.
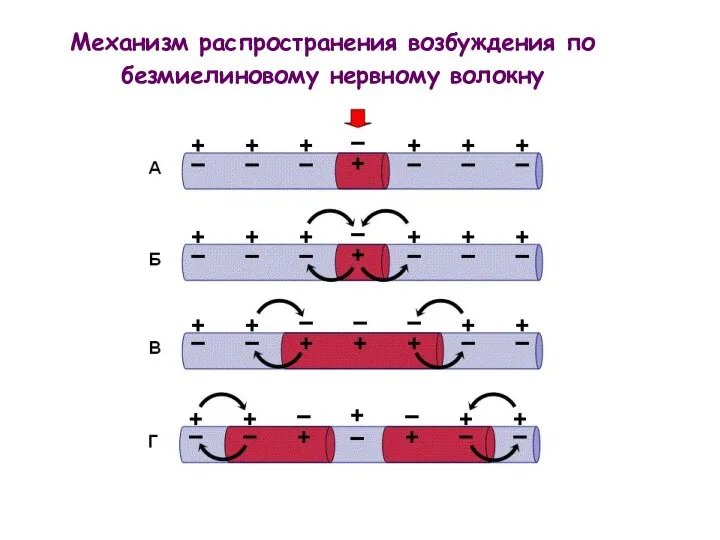
Механизм распространения возбуждения по безмиелиновому нервному волокну
Механизм распространения возбуждения по безмиелиновому нервному волокну
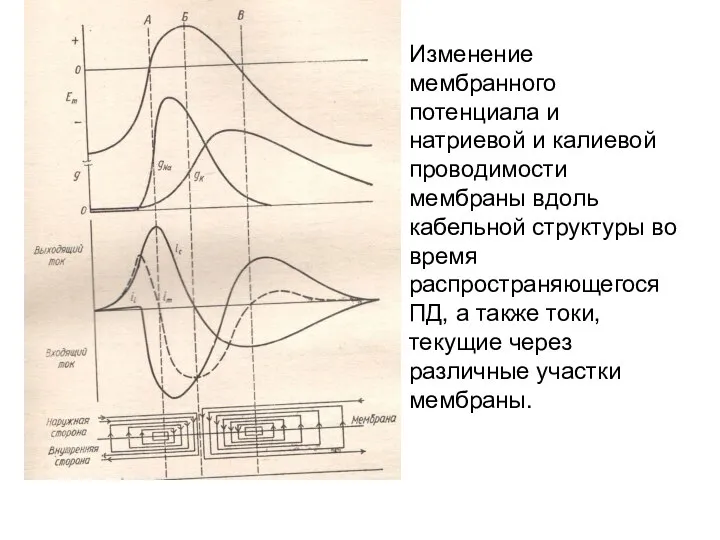
Изменение мембранного потенциала и натриевой и калиевой проводимости мембраны вдоль кабельной
Изменение мембранного потенциала и натриевой и калиевой проводимости мембраны вдоль кабельной
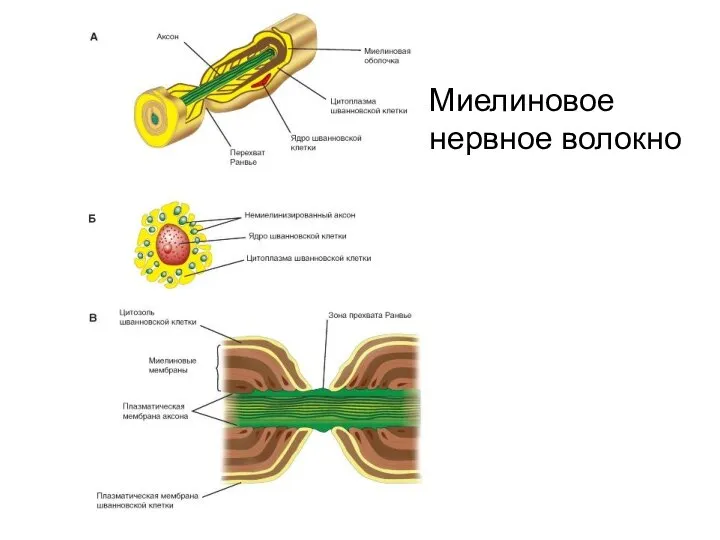
Миелиновое нервное волокно
Миелиновое нервное волокно
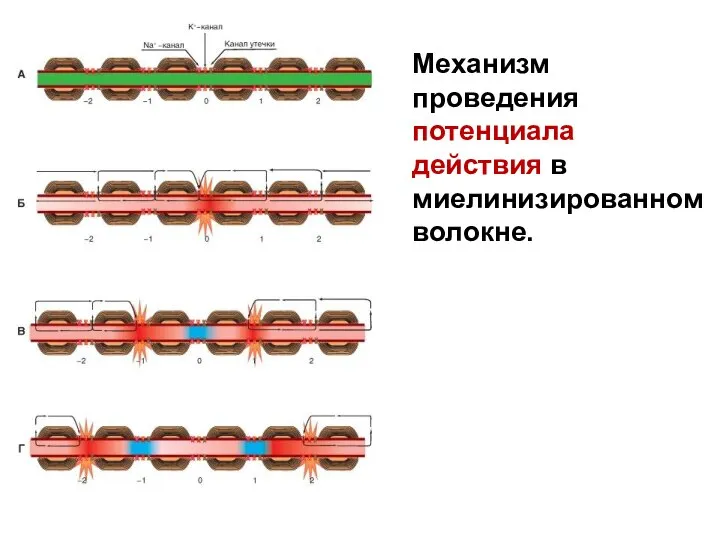
Механизм проведения потенциала действия в миелинизированном волокне.
Механизм проведения потенциала действия в миелинизированном волокне.
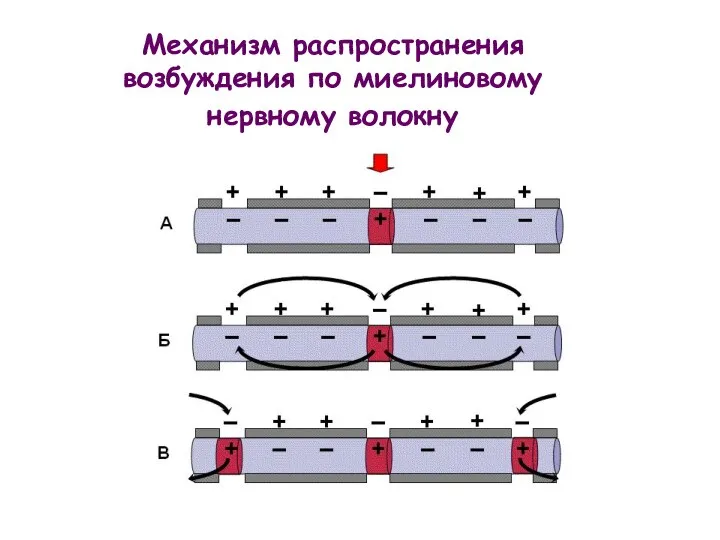
Механизм распространения возбуждения по миелиновому нервному волокну
Механизм распространения возбуждения по миелиновому нервному волокну
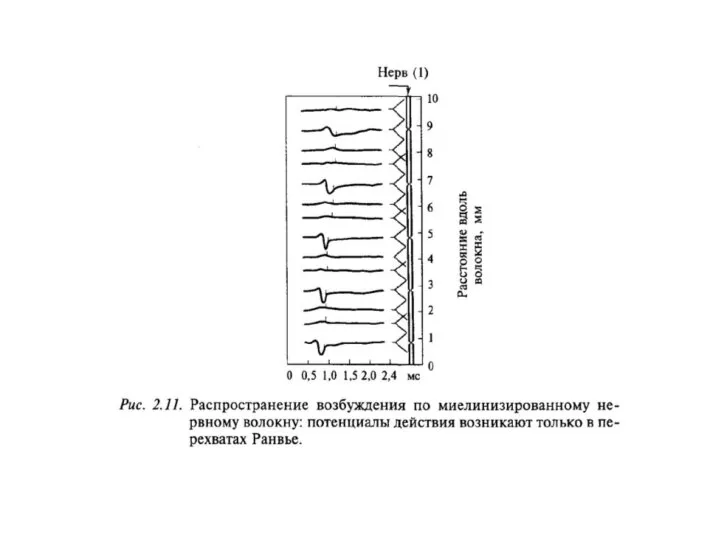
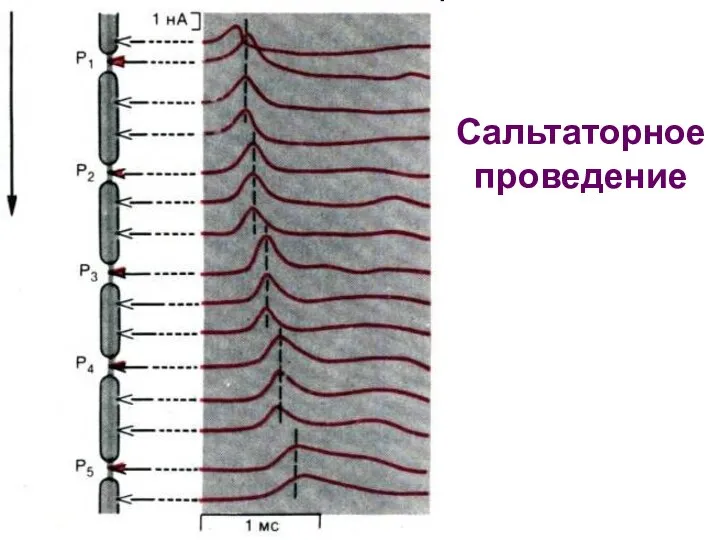
Сальтаторное
проведение
Сальтаторное
проведение
 Пищевые токсикоинфекции. Ботулизм
Пищевые токсикоинфекции. Ботулизм Кинематика
Кинематика Особенности социально-культурной деятельности в театрально-зрелищных учреждениях
Особенности социально-культурной деятельности в театрально-зрелищных учреждениях Schulsachen
Schulsachen Язык разметки XML. Объектно - ориентированное программирование
Язык разметки XML. Объектно - ориентированное программирование Урок 2. Повторение. Обыкновенные дроби
Урок 2. Повторение. Обыкновенные дроби Суперхит на рынке. Изделие №9
Суперхит на рынке. Изделие №9 «Мировое хозяйство и международная торговля.»
«Мировое хозяйство и международная торговля.» Без названия
Без названия Золушка - презентация для начальной школы
Золушка - презентация для начальной школы Сложение гармонических колебаний
Сложение гармонических колебаний  Виды некоммерческого маркетинга Выполнили студенты 4 курса УК Карташева Екатерина Чистякова Надежда
Виды некоммерческого маркетинга Выполнили студенты 4 курса УК Карташева Екатерина Чистякова Надежда  Программирование. ABC Pascal
Программирование. ABC Pascal Двойственный симплекс-метод
Двойственный симплекс-метод Смешинка - 2 - презентация для начальной школы
Смешинка - 2 - презентация для начальной школы Тема 2. Условия принятия управленческих решений условия определенности, риск и неопределенность при принятии управленческих р
Тема 2. Условия принятия управленческих решений условия определенности, риск и неопределенность при принятии управленческих р Лекц10 Выд АФК
Лекц10 Выд АФК Модульная конструкция двигателя Д-436-148 и её влияние на объём и качество технического обслуживания
Модульная конструкция двигателя Д-436-148 и её влияние на объём и качество технического обслуживания Понятие, виды и юридическое значение страхового стажа
Понятие, виды и юридическое значение страхового стажа  National holidays of Kyrgyzstan
National holidays of Kyrgyzstan ТРАДИЦИОННАЯ ПРОГРАММА (под ред. А.Плешакова) «ШКОЛА РОССИИ»
ТРАДИЦИОННАЯ ПРОГРАММА (под ред. А.Плешакова) «ШКОЛА РОССИИ» ГТО («Готов к труду и обороне»)
ГТО («Готов к труду и обороне») Маркетинг - корпоративные стратегии
Маркетинг - корпоративные стратегии Проектирование производства монтажных работ
Проектирование производства монтажных работ Коллектив разработчиков. Лидер
Коллектив разработчиков. Лидер АДРЕСНОЕ ПРОСТРАНСТВО МПС И МЕТОДЫ ДЕШИФРАЦИИ
АДРЕСНОЕ ПРОСТРАНСТВО МПС И МЕТОДЫ ДЕШИФРАЦИИ Общее собрание собственников помещений в многоквартирном доме
Общее собрание собственников помещений в многоквартирном доме Арба парашиет. Четыре главы
Арба парашиет. Четыре главы