- регуляция.ppt

Содержание
- 2. СПОСОБЫ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ I.ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ II.ХИМИЧЕСКАЯ МОДИФИКАЦИЯ БЕЛКОВ III.ИЗМЕНЕНИЕ КОМПАРТМЕНТАЛИЗАЦИИ ВЕЩЕСТВ
- 3. I. ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ РЕГУЛЯЦИЯ НА УРОВНЕ КАТАЛИТИЧЕСКОГО (АКТИВНОГО) ЦЕНТРА АКТИВНОСТЬ ФЕРМЕНТА ОПРЕДЕЛЯЕТСЯ
- 4. ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ 2. РЕГУЛЯЦИЯ НА УРОВНЕ АЛЛОСТЕРИЧЕСКОГО ЦЕНТРА АКТИВНОСТЬ ФЕРМЕНТА РЕГУЛИРУЕТСЯ ЛИГАНДАМИ:
- 5. СИНТЕЗ БЕЛКА НА РИБОСОМЕ В клетках животных многие белки синтезируются в виде молекул—предшественников, требующих модификации для
- 6. II. ХИМИЧЕСКАЯ МОДИФИКАЦИЯ БЕЛКОВ (ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ БЕЛКОВ) ковалентные модификации белка ГИДРОКСИЛИРОВАНИЕ ГЛИКОЗИЛИРОВАНИЕ АЦЕТИЛИРОВАНИЕ МЕТИЛИРОВАНИЕ ФОСФОРИЛИРОВАНИЕ ОГРАНИЧЕННЫЙ
- 7. ГИДРОКСИЛИРОВАНИЕ - присоединение ОН- к определенным остаткам аминокислот ПРИМЕР: коллаген синтезируется в виде проколлагена. Гидроксилирование остатков
- 8. ГЛИКОЗИЛИРОВАНИЕ - присоединение углеводных остатков, образование гликопротеинов Происходит в ЭПС и комплексе Гольджи Перенос олигосахаридов Укорачивание
- 9. АЦЕТИЛИРОВАНИЕ и МЕТИЛИРОВАНИЕ Присоединение ацильной или метильной группы ПРИМЕР: ацетилирование или метилирование гистонов, что влияет на
- 10. ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ ГИСТОНОВ ОКАЗЫВАЕТ ВЛИЯНИЕ НА ТРАНСКРИПЦИОННУЮ АКТИВНОСТЬ ГЕНОВ ВОЗМОЖНЫЙ МЕХАНИЗМ: ИЗМЕНЕНИЕ ВЕЛИЧИНЫ ПОЛОЖИТЕЛЬНОГО ЗАРЯДА ГИСТОНОВ,
- 11. ФОСФОРИЛИРОВАНИЕ Присоединение фосфатной группы. Обратимый процесс. Фосфорилируется остаток серина (треонина) или тирозина. Несмотря на большое количество
- 12. ФОСФОРИЛИРОВАНИЕ КИНАЗА ФОСФОРИЛИРУЕТ ФОСФАТАЗА ДЕФОСФОРИЛИРУЕТ
- 14. ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ – избирательное расщепление белка на фрагменты при участии протеаз Многие ферменты производятся в виде
- 15. ПРИМЕР: АКТИВАЦИЯ ТРИПСИНОГЕНА
- 16. Инсулин синтезируется в виде проинсулина и представляет собой одноцепочечную молекулу. После удаления специфическими протеазами полипептидного сегмента
- 17. ВНУТРИКЛЕТОЧНАЯ Са 2+ -ЗАВИСИМАЯ ПРОТЕАЗА КАЛЬПАИН Нарушение цитоскелета Повреждение мембраны ПОДВЕРГАЕТ ПРОТЕОЛИЗУ МНОГИЕ ВНУТРИКЛЕТОЧНЫЕ БЕЛКИ: БЕЛКИ
- 18. III. ИЗМЕНЕНИЕ КОМПАРТМЕНТАЛИЗАЦИИ ВЕЩЕСТВ В КЛЕТКЕ Ферменты и субстраты разделены мембраной, изменение ее проницаемости для субстратов,
- 19. IV.ТОПОДИНАМИЧЕСКАЯ РЕГУЛЯЦИЯ Основана на динамической неоднородности распределения белков в мембране, их способности образовывать ассоциации.
- 20. I, III, IV – белковые комплексы II – сукцинатдегид-рогеназа V – АТФ-синтаза ПРИМЕР: дыхательная цепь в
- 21. V.ЭКСПРЕССИЯ ГЕНОМА РЕГУЛЯЦИЯ СОДЕРЖАНИЯ БЕЛКА В КЛЕТКЕ ПОСРЕДСТВОМ ИНДУКЦИИ И РЕПРЕССИИ СИНТЕЗА, Т.Е. ИЗМЕНЕНИЕМ СКОРОСТИ ТРАНСКРИПЦИИ
- 22. РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА У ПРОКАРИОТ ИНДУКТОРОМ СИНТЕЗА СЛУЖИТ СУБСТРАТ
- 23. Во всех эукариотических клетках экспрессия генов контролируется регуляторными белками, которые связываются с определенными участками ДНК и
- 24. КОНТРОЛЬ НА УРОВНЕ ТРАНСКРИПЦИИ РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА У ЭУКАРИОТ ИНДУКТОРЫ СИНТЕЗА – СТЕРОИДНЫЕ ГОРМОНЫ
- 25. VI. РЕГУЛЯТОРНЫЕ СИСТЕМЫ С УЧАСТИЕМ ВТОРИЧНЫХ ПОСРЕДНИКОВ КОМПОНЕНТЫ СИСТЕМЫ ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ РЕЦЕПТОР ПРЕОБРАЗОВАТЕЛЬ СИГНАЛА – G
- 26. СПОСОБЫ МЕЖКЛЕТОЧНОЙ КОММУНИКАЦИИ СПОСОБЫ ДОСТАВКИ СИГНАЛЬНЫХ МОЛЕКУЛ К КЛЕТКАМ
- 27. МЕЖКЛЕТОЧНЫЕ СИГНАЛЬНЫЕ МОЛЕКУЛЫ (ПЕРВИЧНЫЕ ПОСРЕДНИКИ) НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ ДИФФУНДИРУЮТ ЧЕРЕЗ МЕМБРАНУ И СВЯЗЫВАЮТСЯ С ВНУТРИКЛЕТОЧНЫМИ РЕЦЕПТОРАМИ
- 28. 1. НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ Стероидные гормоны, вырабатываемые половыми железами и корой надпочечников Тиреоидные гормоны Первичные посредники
- 29. СХЕМА ДЕЙСТВИЯ СТЕРОИДНЫХ И ТИРЕОИДНЫХ ГОРМОНОВ внутриклеточные рецепторы имеют гормон-связывающий домен ДНК-связывающий домен
- 31. МЕХАНИЗМ ДЕЙСТВИЯ ЛИПОФИЛЬНЫХ ГОРМОНОВ, ПРОНИКАЮЩИХ ВНУТРЬ КЛЕТКИ
- 32. 2. ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ Производные арахидоновой кислоты – ЭЙКОЗАНОИДЫ: ПРОСТАГЛАНДИНЫ ЛЕЙКОТРИЕНЫ ПРОСТАЦИКЛИНЫ ТРОМБОКСАНЫ Первичные посредники
- 33. ЭЙКОЗАНОИДЫ ОБРАЗУЮТСЯ ПРАКТИЧЕСКИ ВО ВСЕХ КЛЕТКАХ, ИХ БИОСИНТЕЗ ИНИЦИИРУЕТСЯ ФОСФОЛИПАЗОЙ А2 ДЕЙСТВУЮТ ЧЕРЕЗ МЕМБРАННЫЕ РЕЦЕПТОРЫ, ОСУЩЕСТВЛЯЮТ
- 34. 3. ГИДРОФИЛЬНЫЕ МОЛЕКУЛЫ Первичные посредники ГИДРОФИЛЬНЫЕ ГОРМОНЫ НЕЙРОТРАНСМИТТЕРЫ ФАКТОРЫ РОСТА (эпидермальный фактор роста, фактор роста нейронов,
- 35. СХЕМА ДЕЙСТВИЯ НЕЙРОМЕДИАТОРОВ Первичные посредники
- 36. СХЕМА ДЕЙСТВИЯ ГИДРОФИЛЬНЫХ АГОНИСТОВ Первичные посредники Гидрофильные гормоны Инсулин, факторы роста
- 37. РЕЦЕПТОРЫ И МЕХАНИЗМЫ ИХ ДЕЙСТВИЯ
- 38. РЕЦЕПТОР –СПЕЦИФИЧЕСКИЙ БЕЛОК, КОТОРЫЙ СВЯЗЫВАЕТ СИГНАЛЬНОЕ ВЕЩЕСТВО, ЧТО ПРИВОДИТ В КОНЕЧНОМ ИТОГЕ К РАЗВИТИЮ КЛЕТОЧНОЙ РЕАКЦИИ.
- 39. ХАРАКТЕРИСТИКИ РЕЦЕПТОРОВ
- 40. СЕЛЕКТИВНОСТЬ СВОЙСТВО, ОСНОВАННОЕ НА СТРОГОЙ СТРУКТУРНОЙ СПЕЦИФИЧНОСТИ. ДАННЫЙ ЛИГАНД ДОЛЖЕН БЫТЬ ЛИБО ЕДИНСТВЕННЫМ ВЕЩЕСТВОМ, СВЯЗЫВАЮЩИМСЯ С
- 42. НАСЫЩАЕМОСТЬ ЧИСЛО МЕСТ СВЯЗЫВАНИЯ С ЛИГАНДОМ ДОЛЖНО БЫТЬ КОНЕЧНЫМ.
- 43. СРОДСТВО К ЛИГАНДУ НАСЫЩЕНИЕ РЕЦЕПТОРА ПРОИСХОДИТ ПРИ ФИЗИОЛОГИЧЕСКИХ КОНЦЕНТРАЦИЯХ ЛИГАНДА АКТГ — 0-50 пг/мл Тироксин общий
- 45. Скачать презентацию

СПОСОБЫ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ
I.ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ
II.ХИМИЧЕСКАЯ МОДИФИКАЦИЯ БЕЛКОВ
III.ИЗМЕНЕНИЕ
СПОСОБЫ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ
I.ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ
II.ХИМИЧЕСКАЯ МОДИФИКАЦИЯ БЕЛКОВ
III.ИЗМЕНЕНИЕ
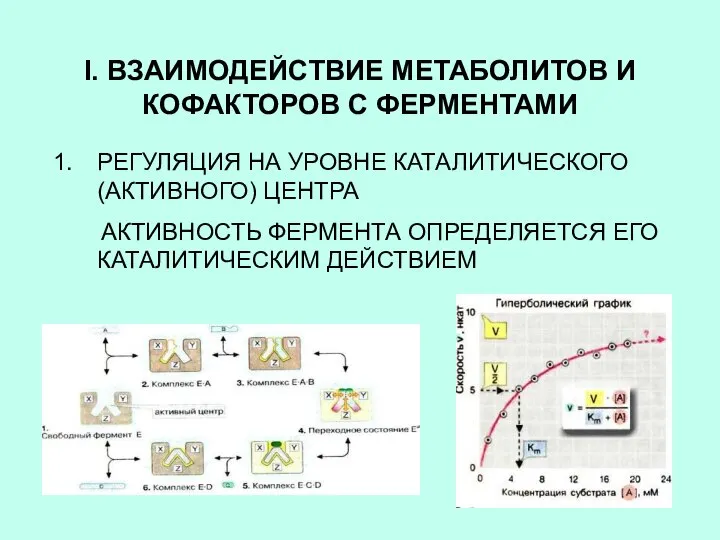
I. ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ
РЕГУЛЯЦИЯ НА УРОВНЕ КАТАЛИТИЧЕСКОГО (АКТИВНОГО)
I. ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ
РЕГУЛЯЦИЯ НА УРОВНЕ КАТАЛИТИЧЕСКОГО (АКТИВНОГО)
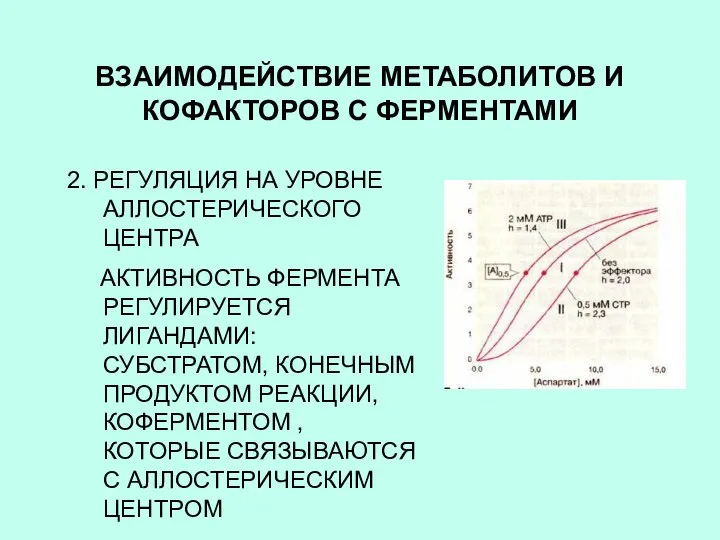
ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ
2. РЕГУЛЯЦИЯ НА УРОВНЕ АЛЛОСТЕРИЧЕСКОГО ЦЕНТРА
ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ
2. РЕГУЛЯЦИЯ НА УРОВНЕ АЛЛОСТЕРИЧЕСКОГО ЦЕНТРА
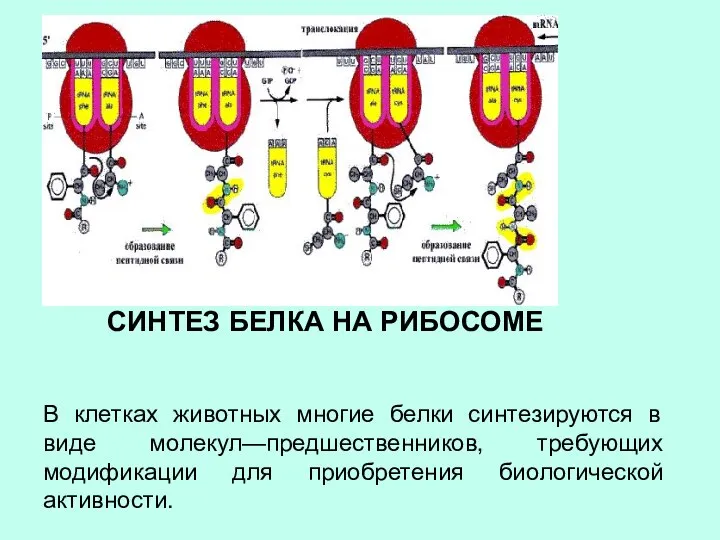
СИНТЕЗ БЕЛКА НА РИБОСОМЕ
В клетках животных многие белки синтезируются в виде
СИНТЕЗ БЕЛКА НА РИБОСОМЕ
В клетках животных многие белки синтезируются в виде

II. ХИМИЧЕСКАЯ МОДИФИКАЦИЯ БЕЛКОВ
(ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ БЕЛКОВ)
ковалентные модификации белка
ГИДРОКСИЛИРОВАНИЕ
ГЛИКОЗИЛИРОВАНИЕ
АЦЕТИЛИРОВАНИЕ
МЕТИЛИРОВАНИЕ
ФОСФОРИЛИРОВАНИЕ
ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ
II. ХИМИЧЕСКАЯ МОДИФИКАЦИЯ БЕЛКОВ
(ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ БЕЛКОВ)
ковалентные модификации белка
ГИДРОКСИЛИРОВАНИЕ
ГЛИКОЗИЛИРОВАНИЕ
АЦЕТИЛИРОВАНИЕ
МЕТИЛИРОВАНИЕ
ФОСФОРИЛИРОВАНИЕ
ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ

ГИДРОКСИЛИРОВАНИЕ - присоединение ОН- к определенным остаткам аминокислот
ПРИМЕР: коллаген синтезируется
ГИДРОКСИЛИРОВАНИЕ - присоединение ОН- к определенным остаткам аминокислот
ПРИМЕР: коллаген синтезируется
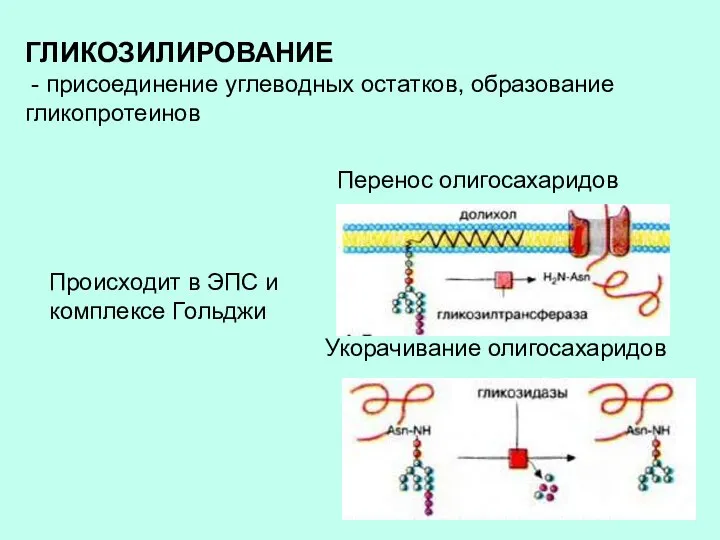
ГЛИКОЗИЛИРОВАНИЕ
- присоединение углеводных остатков, образование гликопротеинов
Происходит в ЭПС и комплексе
ГЛИКОЗИЛИРОВАНИЕ
- присоединение углеводных остатков, образование гликопротеинов
Происходит в ЭПС и комплексе
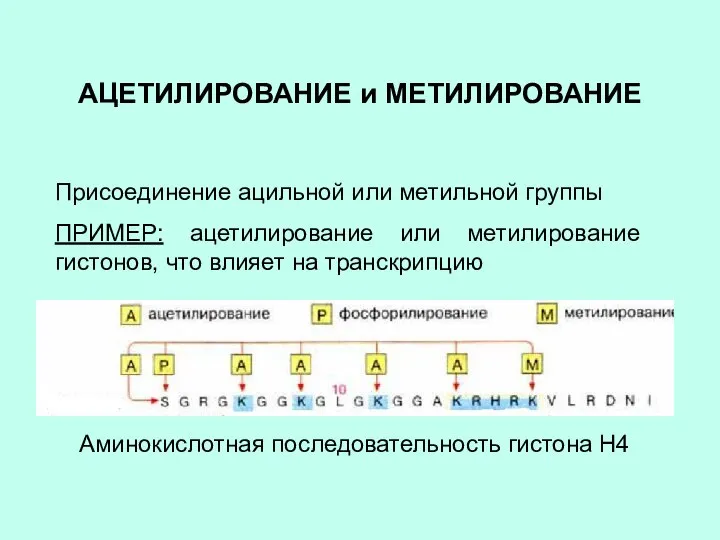
АЦЕТИЛИРОВАНИЕ и МЕТИЛИРОВАНИЕ
Присоединение ацильной или метильной группы
ПРИМЕР: ацетилирование или метилирование гистонов,
АЦЕТИЛИРОВАНИЕ и МЕТИЛИРОВАНИЕ
Присоединение ацильной или метильной группы
ПРИМЕР: ацетилирование или метилирование гистонов,
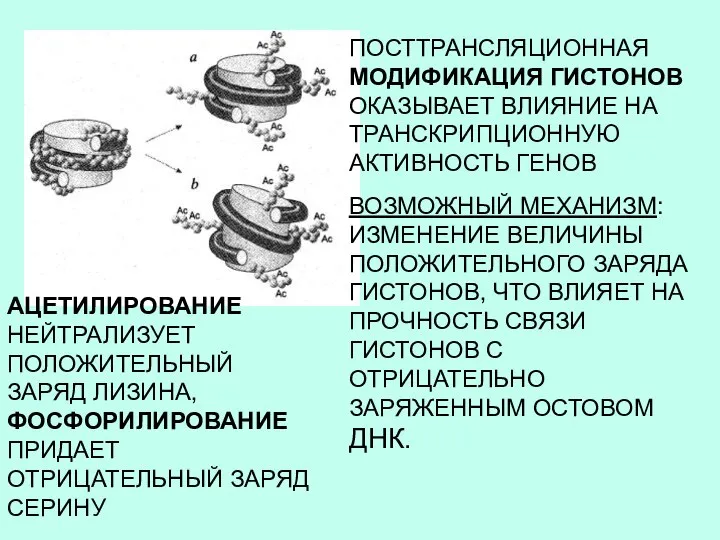
ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ ГИСТОНОВ ОКАЗЫВАЕТ ВЛИЯНИЕ НА ТРАНСКРИПЦИОННУЮ АКТИВНОСТЬ ГЕНОВ
ВОЗМОЖНЫЙ МЕХАНИЗМ: ИЗМЕНЕНИЕ
ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ ГИСТОНОВ ОКАЗЫВАЕТ ВЛИЯНИЕ НА ТРАНСКРИПЦИОННУЮ АКТИВНОСТЬ ГЕНОВ
ВОЗМОЖНЫЙ МЕХАНИЗМ: ИЗМЕНЕНИЕ

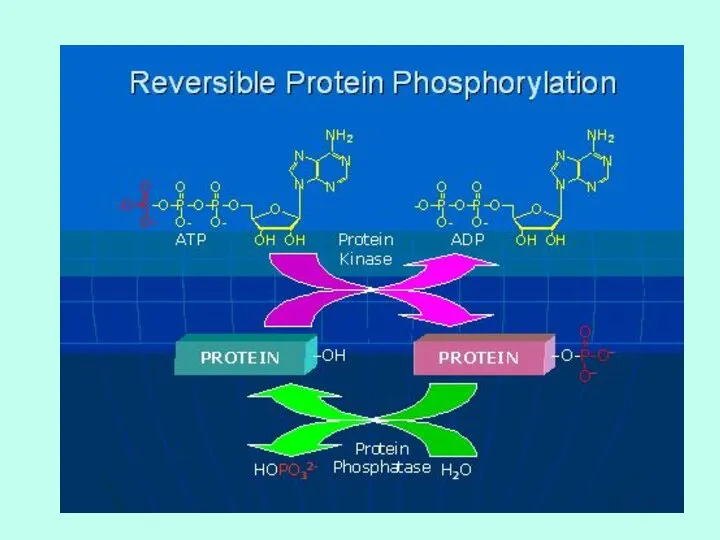
ФОСФОРИЛИРОВАНИЕ
Присоединение фосфатной группы.
Обратимый процесс.
Фосфорилируется остаток серина (треонина) или тирозина.
Несмотря на
ФОСФОРИЛИРОВАНИЕ
Присоединение фосфатной группы.
Обратимый процесс.
Фосфорилируется остаток серина (треонина) или тирозина.
Несмотря на
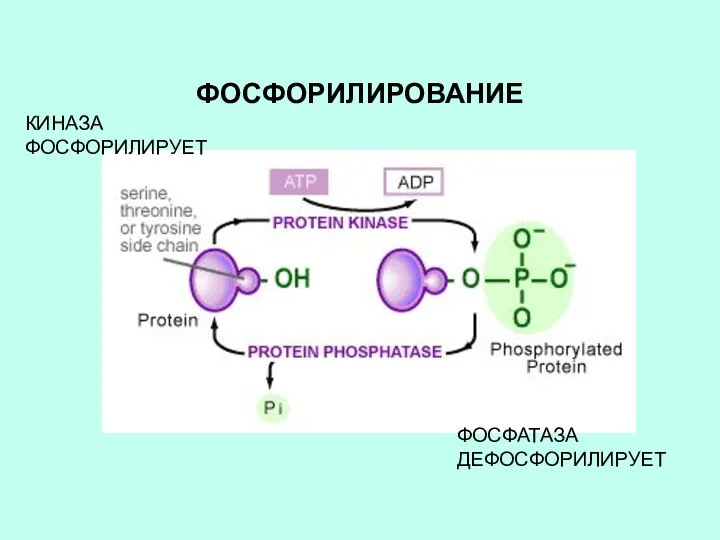
ФОСФОРИЛИРОВАНИЕ
КИНАЗА ФОСФОРИЛИРУЕТ
ФОСФАТАЗА ДЕФОСФОРИЛИРУЕТ
ФОСФОРИЛИРОВАНИЕ
КИНАЗА ФОСФОРИЛИРУЕТ
ФОСФАТАЗА ДЕФОСФОРИЛИРУЕТ
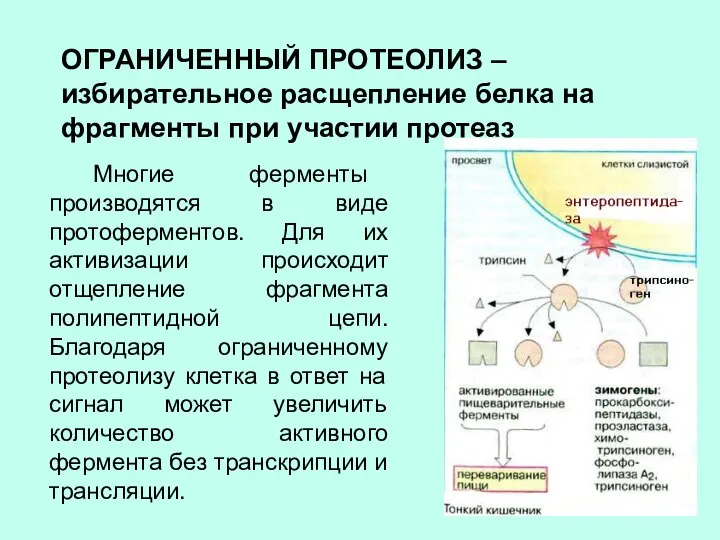
ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ – избирательное расщепление белка на фрагменты при участии протеаз
ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ – избирательное расщепление белка на фрагменты при участии протеаз
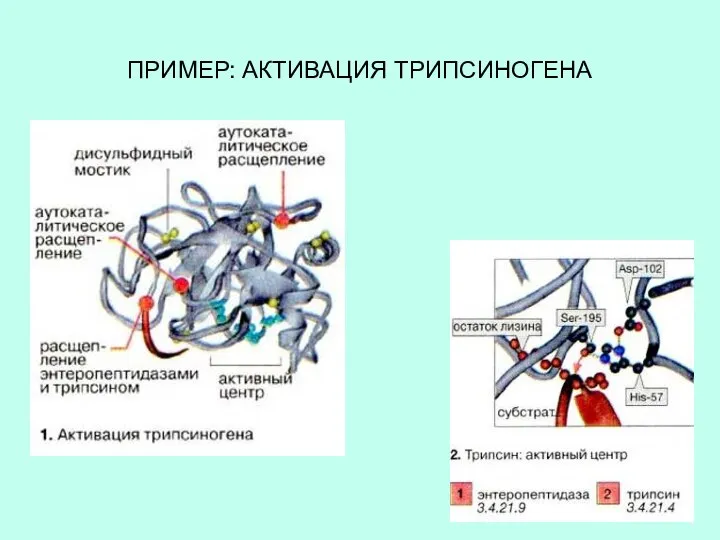
ПРИМЕР: АКТИВАЦИЯ ТРИПСИНОГЕНА
ПРИМЕР: АКТИВАЦИЯ ТРИПСИНОГЕНА
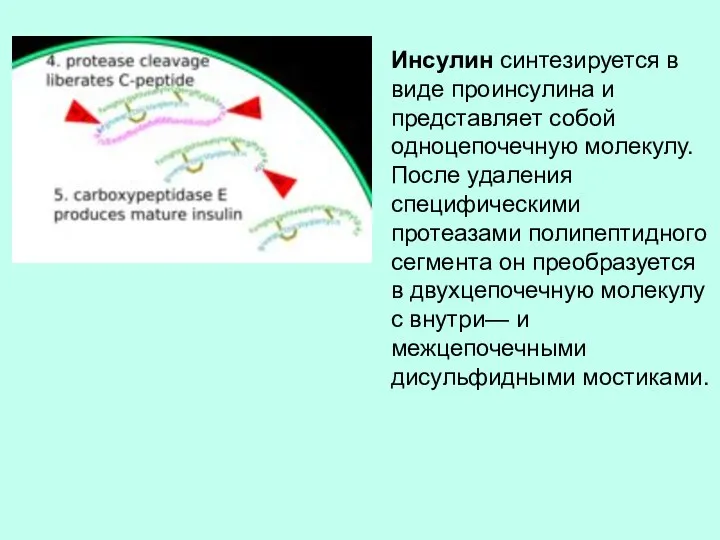
Инсулин синтезируется в виде проинсулина и представляет собой одноцепочечную молекулу. После
Инсулин синтезируется в виде проинсулина и представляет собой одноцепочечную молекулу. После
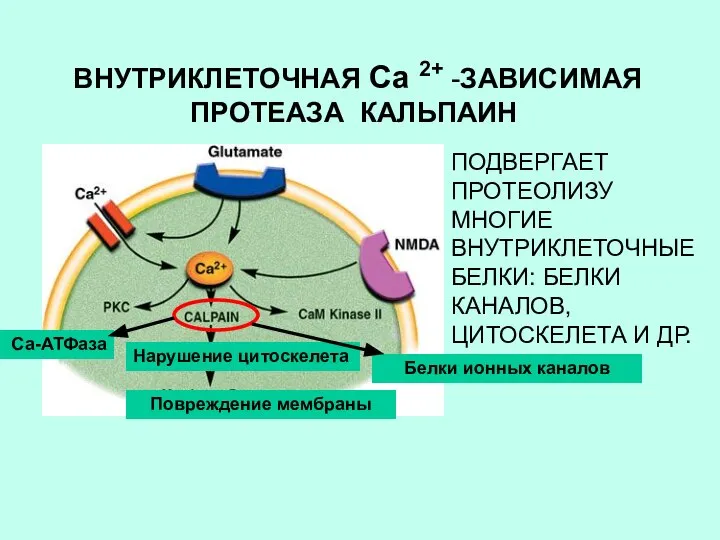
ВНУТРИКЛЕТОЧНАЯ Са 2+ -ЗАВИСИМАЯ ПРОТЕАЗА КАЛЬПАИН
Нарушение цитоскелета
Повреждение мембраны
ПОДВЕРГАЕТ ПРОТЕОЛИЗУ МНОГИЕ
ВНУТРИКЛЕТОЧНАЯ Са 2+ -ЗАВИСИМАЯ ПРОТЕАЗА КАЛЬПАИН
Нарушение цитоскелета
Повреждение мембраны
ПОДВЕРГАЕТ ПРОТЕОЛИЗУ МНОГИЕ
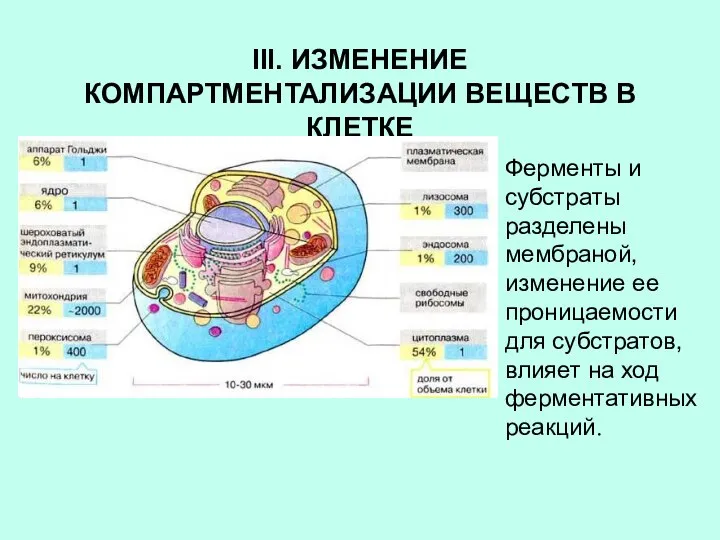
III. ИЗМЕНЕНИЕ КОМПАРТМЕНТАЛИЗАЦИИ ВЕЩЕСТВ В КЛЕТКЕ
Ферменты и субстраты разделены мембраной, изменение
III. ИЗМЕНЕНИЕ КОМПАРТМЕНТАЛИЗАЦИИ ВЕЩЕСТВ В КЛЕТКЕ
Ферменты и субстраты разделены мембраной, изменение
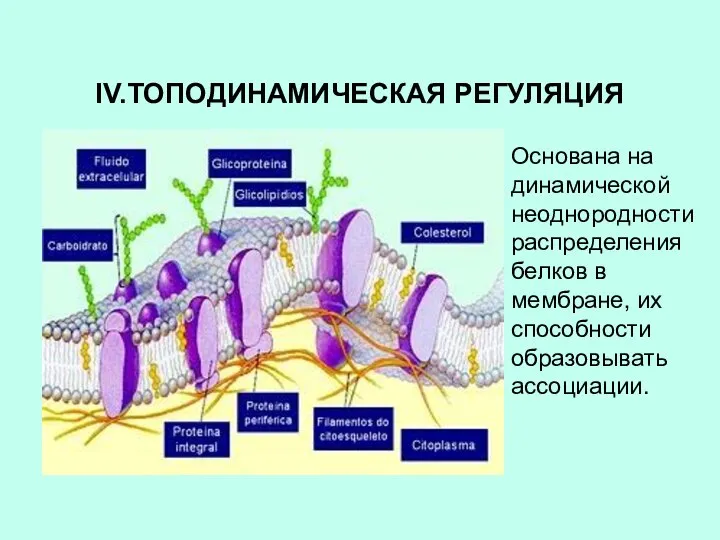
IV.ТОПОДИНАМИЧЕСКАЯ РЕГУЛЯЦИЯ
Основана на динамической неоднородности распределения белков в мембране, их способности
IV.ТОПОДИНАМИЧЕСКАЯ РЕГУЛЯЦИЯ
Основана на динамической неоднородности распределения белков в мембране, их способности
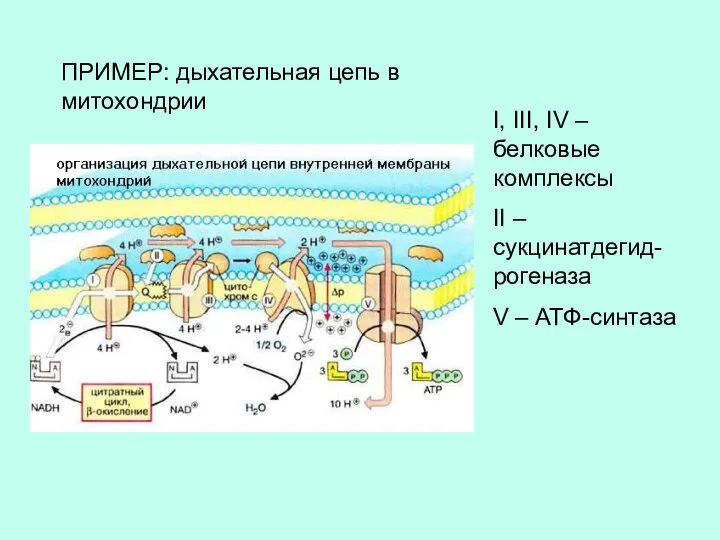
I, III, IV – белковые комплексы
II – сукцинатдегид-рогеназа
V – АТФ-синтаза
ПРИМЕР: дыхательная
I, III, IV – белковые комплексы
II – сукцинатдегид-рогеназа
V – АТФ-синтаза
ПРИМЕР: дыхательная
V.ЭКСПРЕССИЯ ГЕНОМА
РЕГУЛЯЦИЯ СОДЕРЖАНИЯ БЕЛКА В КЛЕТКЕ ПОСРЕДСТВОМ ИНДУКЦИИ И РЕПРЕССИИ СИНТЕЗА,
V.ЭКСПРЕССИЯ ГЕНОМА
РЕГУЛЯЦИЯ СОДЕРЖАНИЯ БЕЛКА В КЛЕТКЕ ПОСРЕДСТВОМ ИНДУКЦИИ И РЕПРЕССИИ СИНТЕЗА,
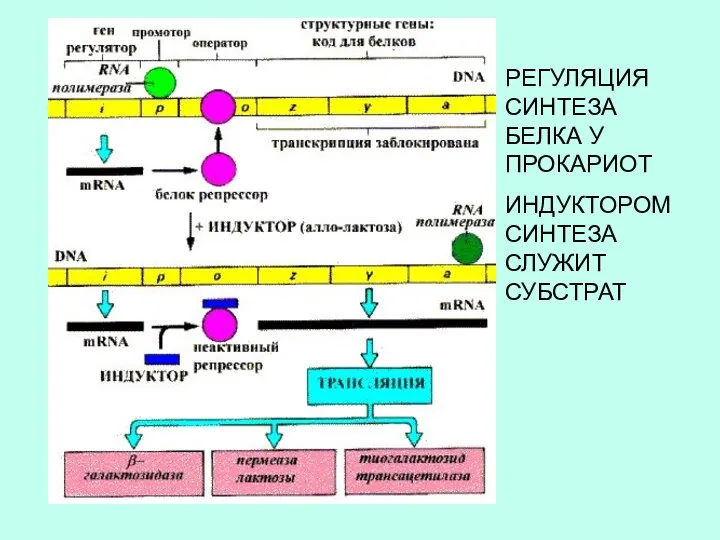
РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА У ПРОКАРИОТ
ИНДУКТОРОМ СИНТЕЗА СЛУЖИТ СУБСТРАТ
РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА У ПРОКАРИОТ
ИНДУКТОРОМ СИНТЕЗА СЛУЖИТ СУБСТРАТ

Во всех эукариотических клетках экспрессия генов контролируется регуляторными белками, которые связываются
Во всех эукариотических клетках экспрессия генов контролируется регуляторными белками, которые связываются
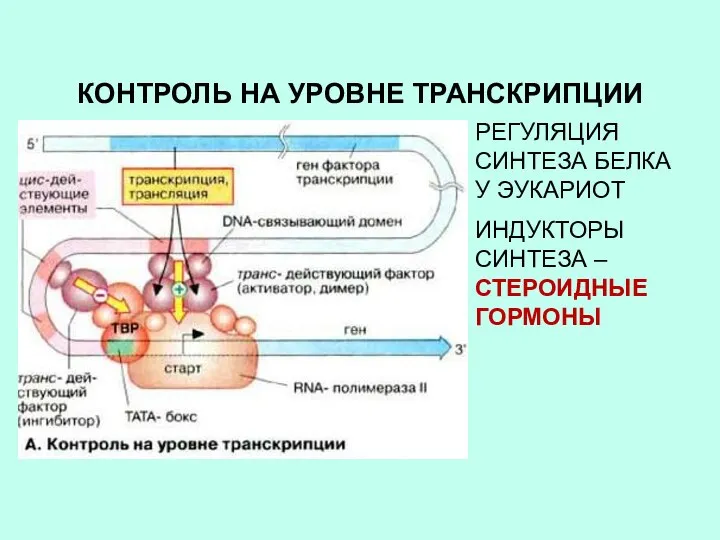
КОНТРОЛЬ НА УРОВНЕ ТРАНСКРИПЦИИ
РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА У ЭУКАРИОТ
ИНДУКТОРЫ СИНТЕЗА – СТЕРОИДНЫЕ
КОНТРОЛЬ НА УРОВНЕ ТРАНСКРИПЦИИ
РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА У ЭУКАРИОТ
ИНДУКТОРЫ СИНТЕЗА – СТЕРОИДНЫЕ
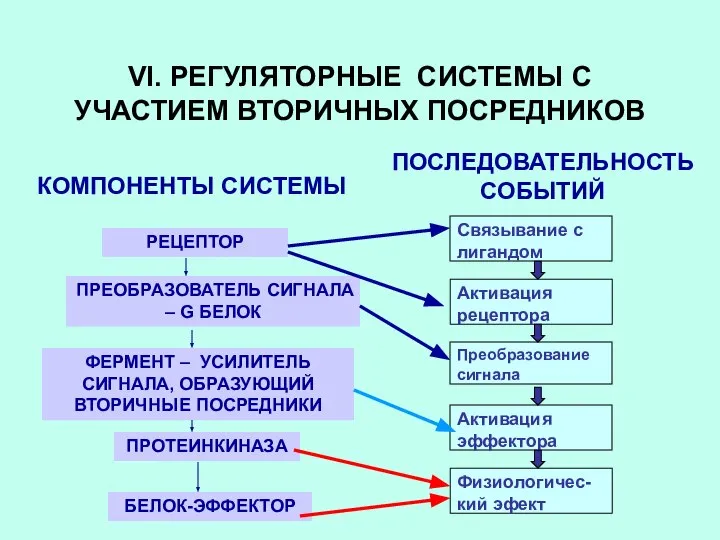
VI. РЕГУЛЯТОРНЫЕ СИСТЕМЫ С УЧАСТИЕМ ВТОРИЧНЫХ ПОСРЕДНИКОВ
КОМПОНЕНТЫ СИСТЕМЫ
ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ
РЕЦЕПТОР
ПРЕОБРАЗОВАТЕЛЬ СИГНАЛА –
VI. РЕГУЛЯТОРНЫЕ СИСТЕМЫ С УЧАСТИЕМ ВТОРИЧНЫХ ПОСРЕДНИКОВ
КОМПОНЕНТЫ СИСТЕМЫ
ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ
РЕЦЕПТОР
ПРЕОБРАЗОВАТЕЛЬ СИГНАЛА –
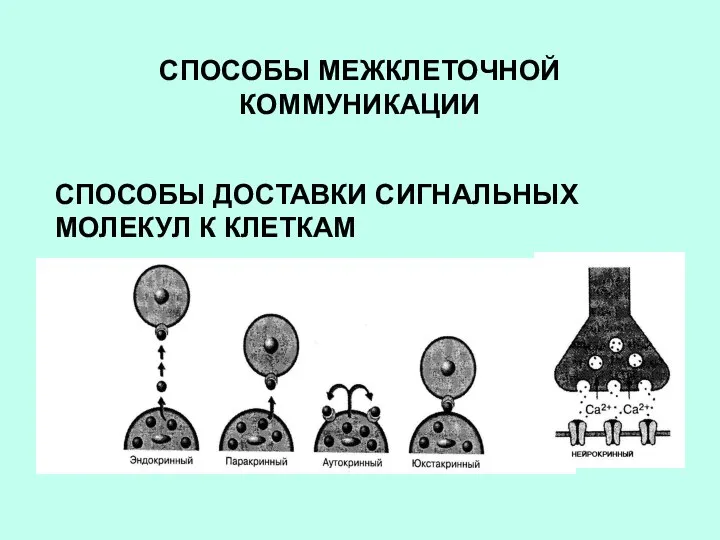
СПОСОБЫ МЕЖКЛЕТОЧНОЙ КОММУНИКАЦИИ
СПОСОБЫ ДОСТАВКИ СИГНАЛЬНЫХ МОЛЕКУЛ К КЛЕТКАМ
СПОСОБЫ МЕЖКЛЕТОЧНОЙ КОММУНИКАЦИИ
СПОСОБЫ ДОСТАВКИ СИГНАЛЬНЫХ МОЛЕКУЛ К КЛЕТКАМ

МЕЖКЛЕТОЧНЫЕ СИГНАЛЬНЫЕ МОЛЕКУЛЫ (ПЕРВИЧНЫЕ ПОСРЕДНИКИ)
НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
ДИФФУНДИРУЮТ ЧЕРЕЗ МЕМБРАНУ И СВЯЗЫВАЮТСЯ
МЕЖКЛЕТОЧНЫЕ СИГНАЛЬНЫЕ МОЛЕКУЛЫ (ПЕРВИЧНЫЕ ПОСРЕДНИКИ)
НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
ДИФФУНДИРУЮТ ЧЕРЕЗ МЕМБРАНУ И СВЯЗЫВАЮТСЯ
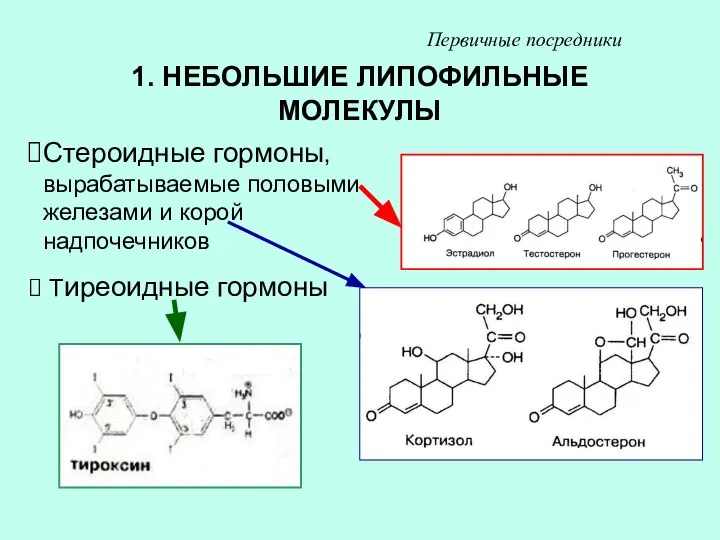
1. НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
Стероидные гормоны, вырабатываемые половыми железами и корой надпочечников
1. НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
Стероидные гормоны, вырабатываемые половыми железами и корой надпочечников
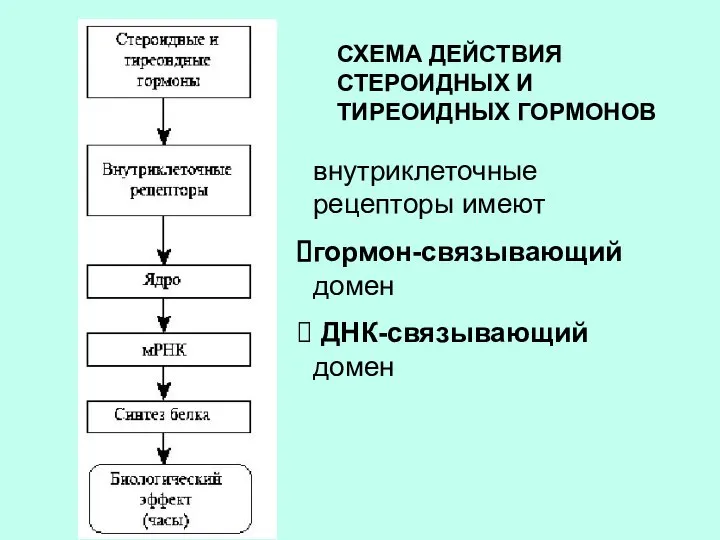
СХЕМА ДЕЙСТВИЯ СТЕРОИДНЫХ И ТИРЕОИДНЫХ ГОРМОНОВ
внутриклеточные рецепторы имеют
гормон-связывающий домен
СХЕМА ДЕЙСТВИЯ СТЕРОИДНЫХ И ТИРЕОИДНЫХ ГОРМОНОВ
внутриклеточные рецепторы имеют
гормон-связывающий домен
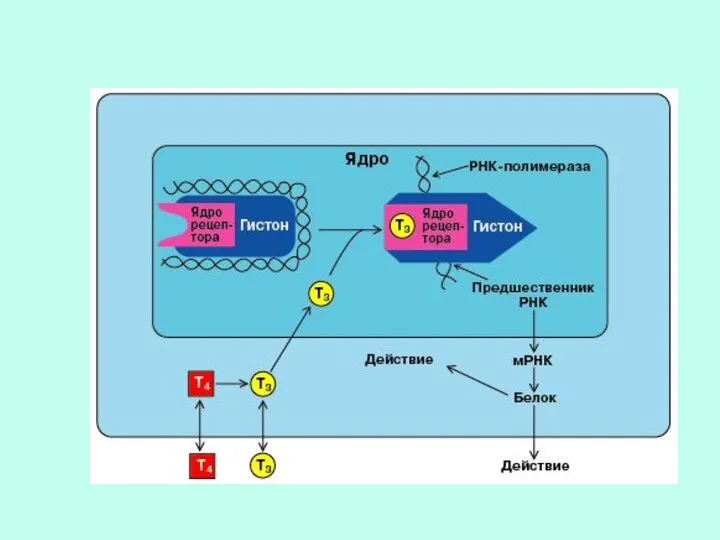

МЕХАНИЗМ ДЕЙСТВИЯ ЛИПОФИЛЬНЫХ ГОРМОНОВ, ПРОНИКАЮЩИХ ВНУТРЬ КЛЕТКИ
МЕХАНИЗМ ДЕЙСТВИЯ ЛИПОФИЛЬНЫХ ГОРМОНОВ, ПРОНИКАЮЩИХ ВНУТРЬ КЛЕТКИ
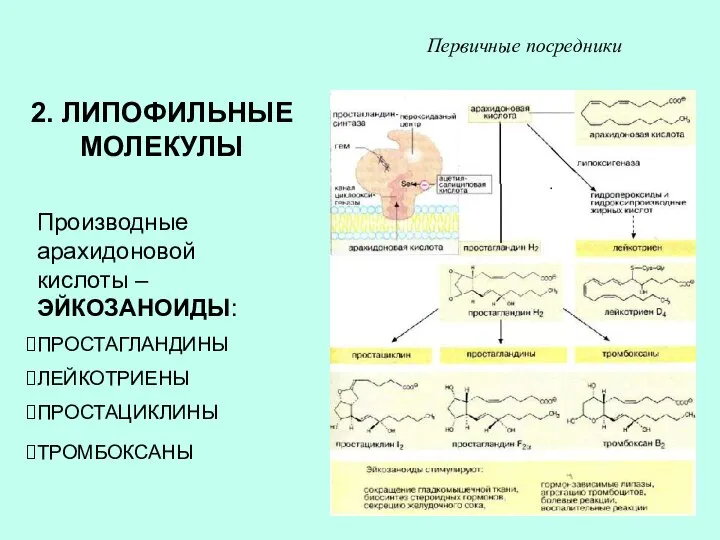
2. ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
Производные арахидоновой кислоты – ЭЙКОЗАНОИДЫ:
ПРОСТАГЛАНДИНЫ
ЛЕЙКОТРИЕНЫ
ПРОСТАЦИКЛИНЫ
ТРОМБОКСАНЫ
Первичные посредники
2. ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ
Производные арахидоновой кислоты – ЭЙКОЗАНОИДЫ:
ПРОСТАГЛАНДИНЫ
ЛЕЙКОТРИЕНЫ
ПРОСТАЦИКЛИНЫ
ТРОМБОКСАНЫ
Первичные посредники

ЭЙКОЗАНОИДЫ ОБРАЗУЮТСЯ ПРАКТИЧЕСКИ ВО ВСЕХ КЛЕТКАХ, ИХ БИОСИНТЕЗ ИНИЦИИРУЕТСЯ ФОСФОЛИПАЗОЙ А2
ЭЙКОЗАНОИДЫ ОБРАЗУЮТСЯ ПРАКТИЧЕСКИ ВО ВСЕХ КЛЕТКАХ, ИХ БИОСИНТЕЗ ИНИЦИИРУЕТСЯ ФОСФОЛИПАЗОЙ А2
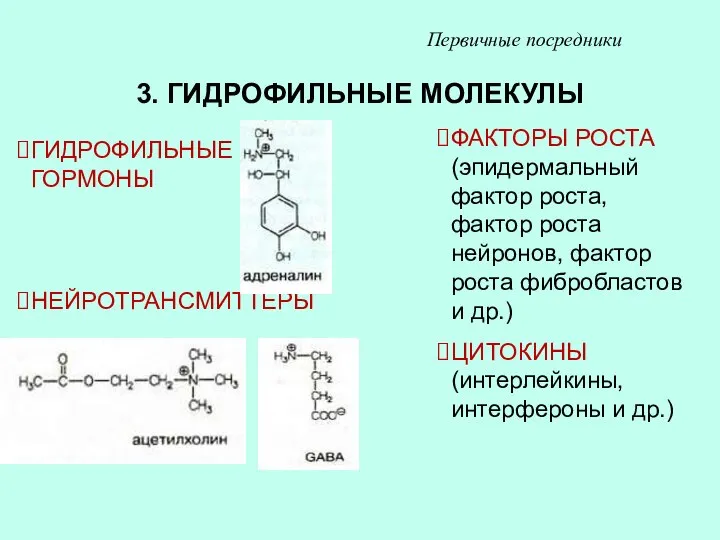
3. ГИДРОФИЛЬНЫЕ МОЛЕКУЛЫ
Первичные посредники
ГИДРОФИЛЬНЫЕ ГОРМОНЫ
НЕЙРОТРАНСМИТТЕРЫ
ФАКТОРЫ РОСТА (эпидермальный фактор роста, фактор роста
3. ГИДРОФИЛЬНЫЕ МОЛЕКУЛЫ
Первичные посредники
ГИДРОФИЛЬНЫЕ ГОРМОНЫ
НЕЙРОТРАНСМИТТЕРЫ
ФАКТОРЫ РОСТА (эпидермальный фактор роста, фактор роста
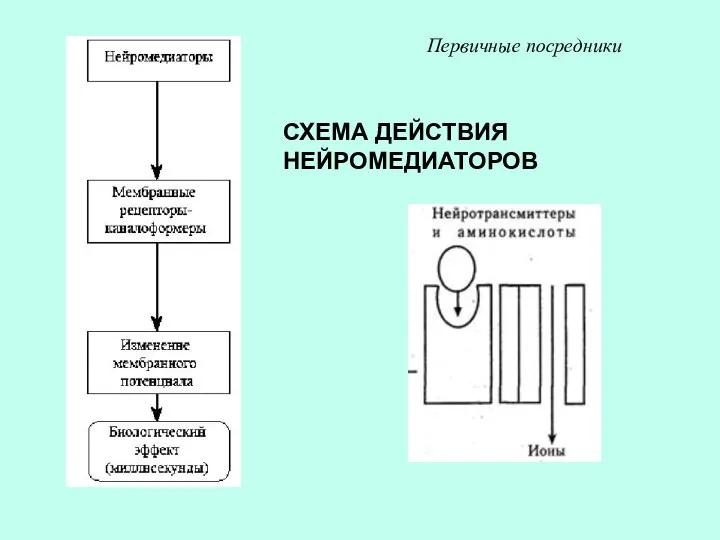
СХЕМА ДЕЙСТВИЯ НЕЙРОМЕДИАТОРОВ
Первичные посредники
СХЕМА ДЕЙСТВИЯ НЕЙРОМЕДИАТОРОВ
Первичные посредники
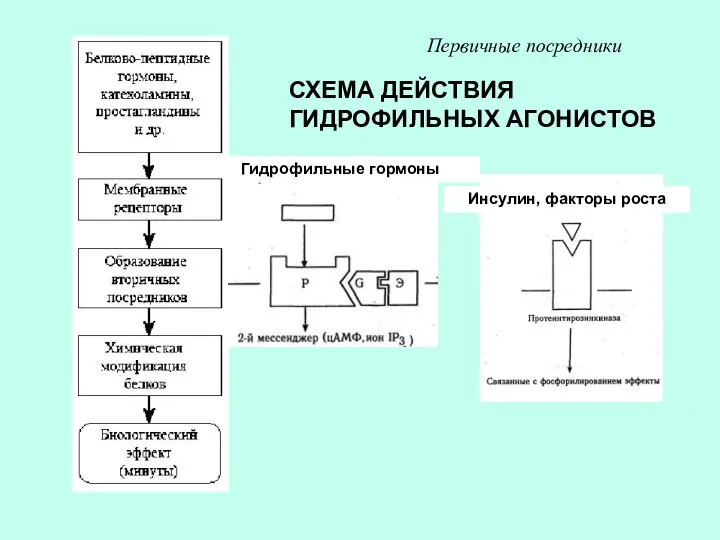
СХЕМА ДЕЙСТВИЯ ГИДРОФИЛЬНЫХ АГОНИСТОВ
Первичные посредники
Гидрофильные гормоны
Инсулин, факторы роста
СХЕМА ДЕЙСТВИЯ ГИДРОФИЛЬНЫХ АГОНИСТОВ
Первичные посредники
Гидрофильные гормоны
Инсулин, факторы роста

РЕЦЕПТОРЫ И МЕХАНИЗМЫ ИХ ДЕЙСТВИЯ
РЕЦЕПТОРЫ И МЕХАНИЗМЫ ИХ ДЕЙСТВИЯ

РЕЦЕПТОР –СПЕЦИФИЧЕСКИЙ БЕЛОК, КОТОРЫЙ СВЯЗЫВАЕТ СИГНАЛЬНОЕ ВЕЩЕСТВО, ЧТО ПРИВОДИТ В КОНЕЧНОМ
РЕЦЕПТОР –СПЕЦИФИЧЕСКИЙ БЕЛОК, КОТОРЫЙ СВЯЗЫВАЕТ СИГНАЛЬНОЕ ВЕЩЕСТВО, ЧТО ПРИВОДИТ В КОНЕЧНОМ

ХАРАКТЕРИСТИКИ РЕЦЕПТОРОВ
ХАРАКТЕРИСТИКИ РЕЦЕПТОРОВ

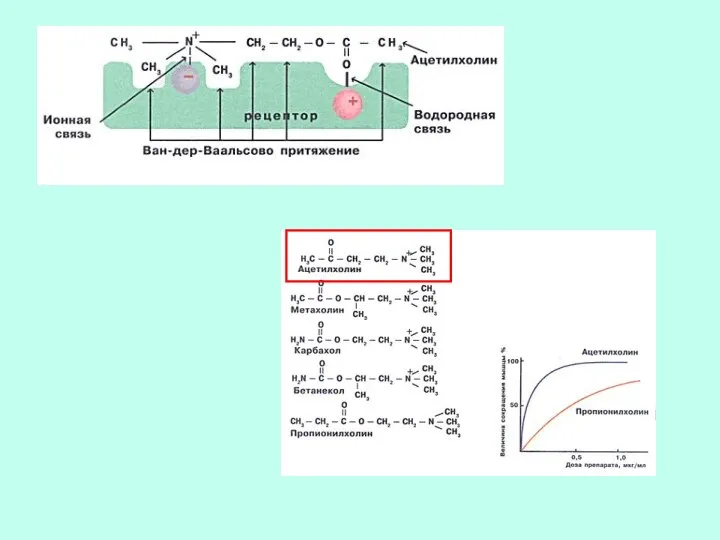
СЕЛЕКТИВНОСТЬ
СВОЙСТВО, ОСНОВАННОЕ НА СТРОГОЙ СТРУКТУРНОЙ СПЕЦИФИЧНОСТИ. ДАННЫЙ ЛИГАНД ДОЛЖЕН БЫТЬ
СЕЛЕКТИВНОСТЬ
СВОЙСТВО, ОСНОВАННОЕ НА СТРОГОЙ СТРУКТУРНОЙ СПЕЦИФИЧНОСТИ. ДАННЫЙ ЛИГАНД ДОЛЖЕН БЫТЬ
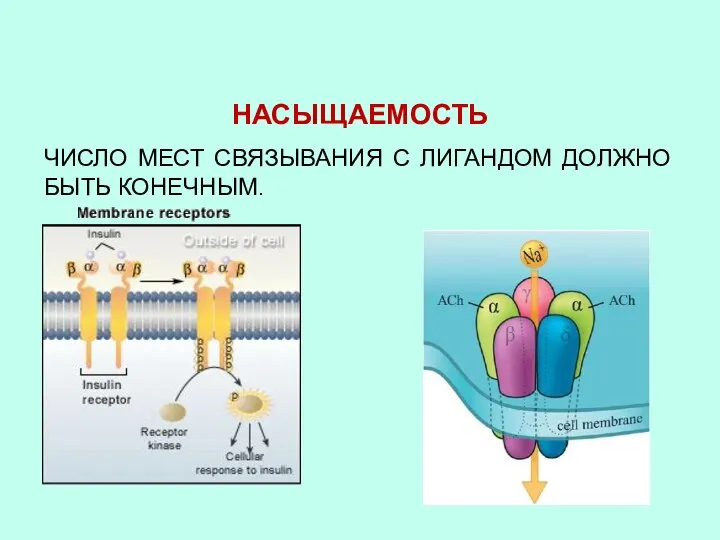
НАСЫЩАЕМОСТЬ
ЧИСЛО МЕСТ СВЯЗЫВАНИЯ С ЛИГАНДОМ ДОЛЖНО БЫТЬ КОНЕЧНЫМ.
НАСЫЩАЕМОСТЬ
ЧИСЛО МЕСТ СВЯЗЫВАНИЯ С ЛИГАНДОМ ДОЛЖНО БЫТЬ КОНЕЧНЫМ.

СРОДСТВО К ЛИГАНДУ
НАСЫЩЕНИЕ РЕЦЕПТОРА ПРОИСХОДИТ ПРИ ФИЗИОЛОГИЧЕСКИХ КОНЦЕНТРАЦИЯХ ЛИГАНДА
АКТГ —
СРОДСТВО К ЛИГАНДУ
НАСЫЩЕНИЕ РЕЦЕПТОРА ПРОИСХОДИТ ПРИ ФИЗИОЛОГИЧЕСКИХ КОНЦЕНТРАЦИЯХ ЛИГАНДА
АКТГ —
 Лимфатическая система, systema lymphatica, - часть кровеносной (венозной) системы
Лимфатическая система, systema lymphatica, - часть кровеносной (венозной) системы Поздравление с 8 марта
Поздравление с 8 марта Технология обследования моторных функций и речи Доктор педагогических наук, профессор Архипова Е.Ф.
Технология обследования моторных функций и речи Доктор педагогических наук, профессор Архипова Е.Ф.  Методы интеллектуального анализа данных Мартин Браун (Martin C. Brown)
Методы интеллектуального анализа данных Мартин Браун (Martin C. Brown) СТЕРИЛИЗАЦИЯ ПИТАТЕЛЬНЫХ СРЕД
СТЕРИЛИЗАЦИЯ ПИТАТЕЛЬНЫХ СРЕД Білково-енергетична недостатність у дітей
Білково-енергетична недостатність у дітей  Мотивация персонала
Мотивация персонала Пособия для обучающихся, как конструктор учебных ситуаций
Пособия для обучающихся, как конструктор учебных ситуаций Творець мій і Господь. Anne Steele (1716 - 1778)
Творець мій і Господь. Anne Steele (1716 - 1778) Анализаторы человека
Анализаторы человека МОДНОЕ ДЕФИЛЕ
МОДНОЕ ДЕФИЛЕ Содержание и психологические факторы эффективной управленческой деятельности в таможне Подготовили: Никул Алла, Шунайлова Жан
Содержание и психологические факторы эффективной управленческой деятельности в таможне Подготовили: Никул Алла, Шунайлова Жан Оценка деятельности совета директоров и вознаграждение членов совета директоров
Оценка деятельности совета директоров и вознаграждение членов совета директоров Построение аксонометрических проекций. (Лекция 9)
Построение аксонометрических проекций. (Лекция 9) Технологическая линия первичной переработки скота
Технологическая линия первичной переработки скота  Протокол об административном правонарушении: понятие , процессуальное значение ,порядок составления Моисеенкова Ольга Митченко
Протокол об административном правонарушении: понятие , процессуальное значение ,порядок составления Моисеенкова Ольга Митченко Викторина о спорте
Викторина о спорте ГБОУ ВПО Саратовский ГМУ им. В.И. Разумовского Минздравсоцразвития России Кафедра инфекционных болезней
ГБОУ ВПО Саратовский ГМУ им. В.И. Разумовского Минздравсоцразвития России Кафедра инфекционных болезней Презентация "Русские прялки" - скачать презентации по МХК
Презентация "Русские прялки" - скачать презентации по МХК Бухгалтерские и юридические услуги, кадровый учет, услуги эколога для ИП, ООО (ОДО), ЧУП
Бухгалтерские и юридические услуги, кадровый учет, услуги эколога для ИП, ООО (ОДО), ЧУП Элементы теории приема и обработки информации
Элементы теории приема и обработки информации Международные экономические отношения 1.Понятие мирового хозяйства и формы МЭО. 2. Глобализация мировой экономики. 3. Протекци
Международные экономические отношения 1.Понятие мирового хозяйства и формы МЭО. 2. Глобализация мировой экономики. 3. Протекци Устройство токарного станка для обработки древесины. (Технический труд. 6 класс)
Устройство токарного станка для обработки древесины. (Технический труд. 6 класс) Методи та системи неруйнівного контролю
Методи та системи неруйнівного контролю Биография Мартина Лютера
Биография Мартина Лютера Экономический рост
Экономический рост Переменные в VB
Переменные в VB Типы запросов и справок
Типы запросов и справок