- Метаболизм липидов. Метаболизм триацилглицеридов. (Лекция 9)

Содержание
- 2. Депонирование нейтрального жира (ТАГ) Жиры и гликоген – формы депонирования энергетического материала. Жиры – наиболее долговременные
- 3. Локализация синтеза ТАГ В печени и жировой ткани (преимущественно, а также в др). В жировой ткани
- 4. Пути образования глицерол -3-фосфата в печени и жировой ткани СН2 - ОН I СН - ОН
- 5. Образование фосфатидной кислоты (последовательная этерификация глицерол-3-фосфата СН2 – ОН СН2 – О – СО – R1
- 6. Источники жирных кислот для синтеза ТАГ Жировая ткань. а) Преимущественно ЖК, освободившиеся при гидролизе жиров ХМ(
- 7. Синтез ВЖК В абсорбтивный период (избыток углеводов). В цитоплазме. Активируется инсулином Субстрат: Ацетил-КоА - метаболит окисления
- 8. Особенности синтеза ВЖК Синтез представлен последовательными реакциями, ведущими к удлинению молекулы ЖК. Катализируются реакции полифункциональным ферментом
- 9. Синтез ТАГ 1. Активация жирной кислоты: R1 СООН + АТФ + НSКоА → R1СО- SКоА +АМФ
- 10. Триацилглицеролы ( жиры), депонированные в адипоцитах в абсорбтивный период, используются как источник энергии в период голодания
- 11. Использование депонированного жира (мобилизация). Тканевой липолиз. Регуляция Тканевой липолиз ( мобилизация тканевого жира в клетках тканей
- 12. Тканевой липолиз сн2-о-со-R1 сн2-он сн2-он сн2-он I I I I cн-о-со-R2 cн-о-со-R2 сн-о-со-R2 сн-оH I I
- 13. Судьба продуктов тканевого липолиза Если тканевой липолиз в жировой ткани,то ВЖК, в большей степени, выходят в
- 14. Регуляция тканевого липолиза
- 15. Источники и значение жирных кислот Источники: - Продукты гидролиза (переваривания) экзогенных жиров; Продукты тканевого липолиза; Синтез
- 16. Бета-окисление жирных кислот. Значение-энергетическое Окисление - в матриксе митохондрий Жирные кислоты в цитоплазме активируются АТФ и
- 17. Бета- окисление (продолжение) Только в аэробных условиях. Окисление происходит по бета-схеме, т.е. окисляется бета-углеродный атом. Цикл
- 18. Бета-окисление высших жирных кислот СН3 (СН)n СН2 –СН2 –СО-SKoA СН3 (СН)n СН = СН –СО-SKoA ФАД
- 19. Окисление глицерина СН2- ОН СН2 - ОН СОН I 1 I 2 I СН - ОН
- 20. Кетоновые тела. Кетогенез. Ацетоуксусная. Бета-гидроксибутират, ацетон Субстрат – Ацетил-КоА Только в митохондрии печени при низком соотношении
- 21. Кетогенез СН3СО-SКоА + СН3СО-SКоА СН3-СО- СН2- СО-SКоА ацетоацетилКоА СН3СО-SКоА СООН-СН2 - СН- СО-SКоА гидрокси- метилглутарил КоА
- 23. Скачать презентацию

Депонирование нейтрального жира (ТАГ)
Жиры и гликоген – формы депонирования энергетического материала.
Жиры
Депонирование нейтрального жира (ТАГ)
Жиры и гликоген – формы депонирования энергетического материала.
Жиры

Локализация синтеза ТАГ
В печени и жировой ткани (преимущественно, а также в
Локализация синтеза ТАГ
В печени и жировой ткани (преимущественно, а также в

Пути образования глицерол -3-фосфата в печени и жировой ткани
СН2 -
Пути образования глицерол -3-фосфата в печени и жировой ткани
СН2 -
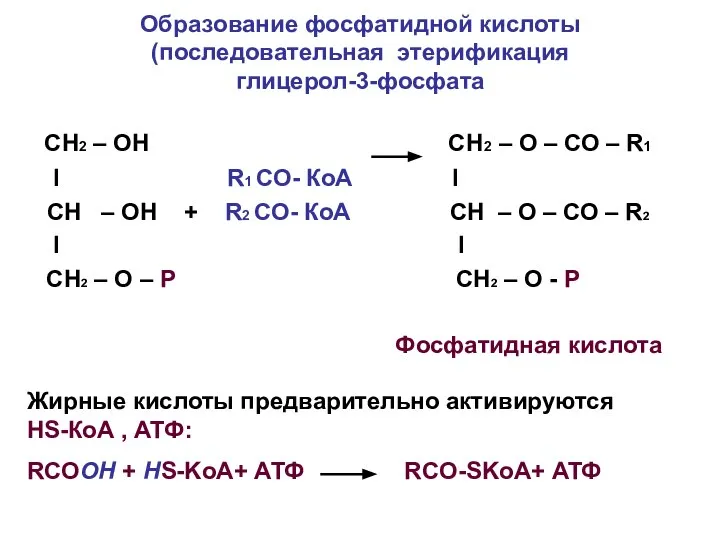
Образование фосфатидной кислоты (последовательная этерификация
глицерол-3-фосфата
СН2 – ОН СН2 –
Образование фосфатидной кислоты (последовательная этерификация
глицерол-3-фосфата
СН2 – ОН СН2 –

Источники жирных кислот для синтеза ТАГ
Жировая ткань.
а) Преимущественно ЖК, освободившиеся
Источники жирных кислот для синтеза ТАГ
Жировая ткань.
а) Преимущественно ЖК, освободившиеся

Синтез ВЖК
В абсорбтивный период (избыток углеводов). В цитоплазме. Активируется инсулином
Субстрат: Ацетил-КоА
Синтез ВЖК
В абсорбтивный период (избыток углеводов). В цитоплазме. Активируется инсулином
Субстрат: Ацетил-КоА

Особенности синтеза ВЖК
Синтез представлен последовательными реакциями, ведущими к удлинению молекулы ЖК.
Особенности синтеза ВЖК
Синтез представлен последовательными реакциями, ведущими к удлинению молекулы ЖК.

Синтез ТАГ
1. Активация жирной кислоты:
R1 СООН + АТФ +
Синтез ТАГ
1. Активация жирной кислоты:
R1 СООН + АТФ +

Триацилглицеролы ( жиры), депонированные в адипоцитах в абсорбтивный период, используются как
Триацилглицеролы ( жиры), депонированные в адипоцитах в абсорбтивный период, используются как

Использование депонированного жира (мобилизация). Тканевой липолиз. Регуляция
Тканевой липолиз ( мобилизация тканевого
Использование депонированного жира (мобилизация). Тканевой липолиз. Регуляция
Тканевой липолиз ( мобилизация тканевого

Тканевой липолиз
сн2-о-со-R1 сн2-он сн2-он сн2-он
I I I I
cн-о-со-R2 cн-о-со-R2 сн-о-со-R2 сн-оH
I
Тканевой липолиз
сн2-о-со-R1 сн2-он сн2-он сн2-он
I I I I
cн-о-со-R2 cн-о-со-R2 сн-о-со-R2 сн-оH
I

Судьба продуктов тканевого липолиза
Если тканевой липолиз в жировой ткани,то ВЖК, в
Судьба продуктов тканевого липолиза
Если тканевой липолиз в жировой ткани,то ВЖК, в
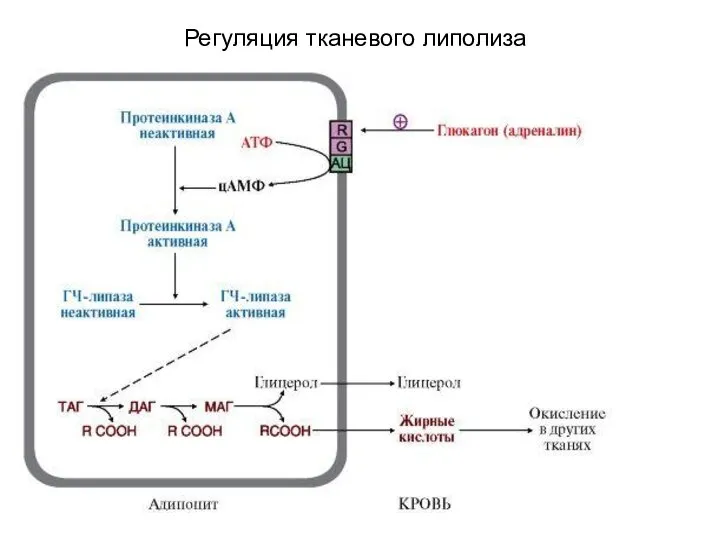
Регуляция тканевого липолиза
Регуляция тканевого липолиза

Источники и значение жирных кислот
Источники:
- Продукты гидролиза (переваривания) экзогенных жиров;
Продукты
Источники и значение жирных кислот
Источники:
- Продукты гидролиза (переваривания) экзогенных жиров;
Продукты

Бета-окисление жирных кислот. Значение-энергетическое
Окисление - в матриксе митохондрий
Жирные кислоты в цитоплазме
Бета-окисление жирных кислот. Значение-энергетическое
Окисление - в матриксе митохондрий
Жирные кислоты в цитоплазме

Бета- окисление (продолжение)
Только в аэробных условиях.
Окисление происходит по бета-схеме, т.е.
Бета- окисление (продолжение)
Только в аэробных условиях.
Окисление происходит по бета-схеме, т.е.
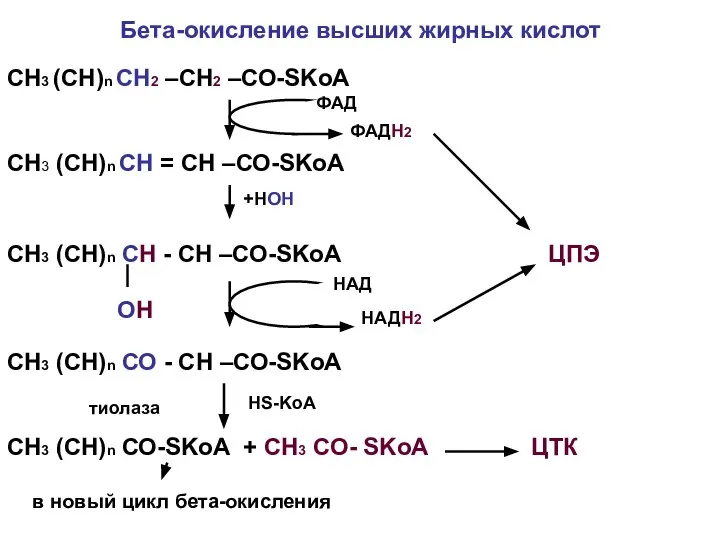
Бета-окисление высших жирных кислот
СН3 (СН)n СН2 –СН2 –СО-SKoA
СН3 (СН)n СН =
Бета-окисление высших жирных кислот
СН3 (СН)n СН2 –СН2 –СО-SKoA
СН3 (СН)n СН =
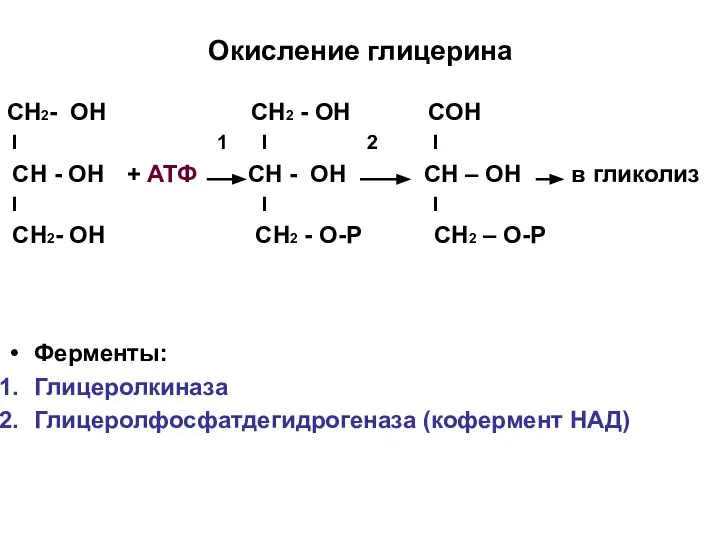
Окисление глицерина
СН2- ОН СН2 - ОН СОН
I 1 I
Окисление глицерина
СН2- ОН СН2 - ОН СОН
I 1 I

Кетоновые тела. Кетогенез.
Ацетоуксусная. Бета-гидроксибутират, ацетон
Субстрат – Ацетил-КоА
Только в митохондрии печени при
Кетоновые тела. Кетогенез.
Ацетоуксусная. Бета-гидроксибутират, ацетон
Субстрат – Ацетил-КоА
Только в митохондрии печени при
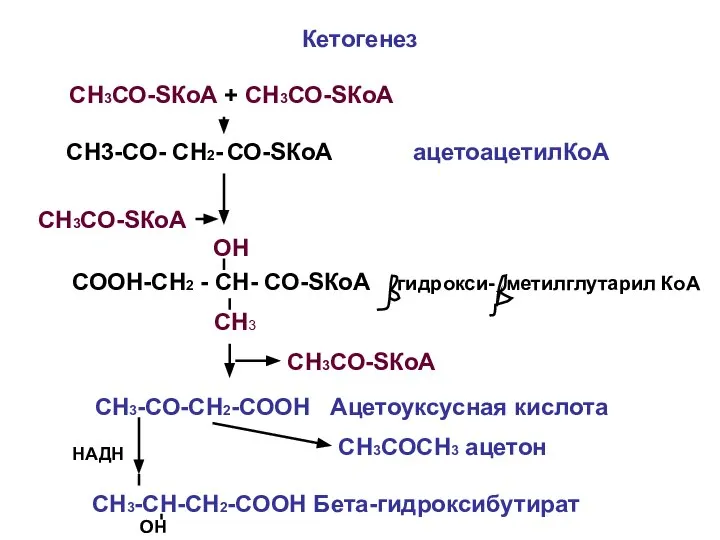
Кетогенез
СН3СО-SКоА + СН3СО-SКоА
СН3-СО- СН2- СО-SКоА ацетоацетилКоА
СН3СО-SКоА
СООН-СН2 - СН- СО-SКоА
Кетогенез
СН3СО-SКоА + СН3СО-SКоА
СН3-СО- СН2- СО-SКоА ацетоацетилКоА
СН3СО-SКоА
СООН-СН2 - СН- СО-SКоА
 Щелочи
Щелочи Элементы группы галогенов
Элементы группы галогенов Агрегирование ПАВ в воде и неполярных растворителях
Агрегирование ПАВ в воде и неполярных растворителях Строение атома
Строение атома Хроматографія. Мікрогетерогенні дисперсні системи
Хроматографія. Мікрогетерогенні дисперсні системи Воспламенение (зажигание) газовых смесей
Воспламенение (зажигание) газовых смесей Излучение и поглощение света атомами. Виды спектров, спектральный анализ
Излучение и поглощение света атомами. Виды спектров, спектральный анализ Дисперсные
Дисперсные Физические свойства минералов
Физические свойства минералов Синтетикалық талшықтар
Синтетикалық талшықтар Характеристика азота как химического элемента и простого вещества. Урок химии в 9 классе.
Характеристика азота как химического элемента и простого вещества. Урок химии в 9 классе. Белки, протеины, полипептиды
Белки, протеины, полипептиды Щелочные металлы
Щелочные металлы Мысты электролитті тазалау
Мысты электролитті тазалау Изготовление мыла в домашних условиях
Изготовление мыла в домашних условиях Химия и пища. Маркировка, упаковка пищевых и гигиенических продуктов, и умение их читать
Химия и пища. Маркировка, упаковка пищевых и гигиенических продуктов, и умение их читать Структура и содержание КИМ-22
Структура и содержание КИМ-22 Кислород
Кислород Гетерофункциональные соединения. (Лекция 14)
Гетерофункциональные соединения. (Лекция 14) Геометрия молекул
Геометрия молекул Электролитическая диссоциация
Электролитическая диссоциация Растворы и их классификация
Растворы и их классификация Хімія і медицина
Хімія і медицина  Основные характеристики ковалентной связи
Основные характеристики ковалентной связи Энергетика и направление химических реакций
Энергетика и направление химических реакций Молекулярно-массовые характеристики высокомолекулярных соединений
Молекулярно-массовые характеристики высокомолекулярных соединений Қазіргі кезде қолданылатын дезинсектицидтер
Қазіргі кезде қолданылатын дезинсектицидтер Типы химических реакций. Класс «оксиды»
Типы химических реакций. Класс «оксиды»