- Пептиды. Белки

Содержание
- 2. Цели лекции: Обучающая - Сформировать знания о строении пептидов и белков и их биологической роли. Развивающая
- 3. План 1. Классификация пептидов и белков. Основные функции белков. 2. Пространственное строение пептидов и белков. 3.
- 4. 1. Классификация пептидов и белков
- 5. Белки и пептиды- полимерные азотсодержащие органические вещества состоящие из остатков аминокислот, соединенные пептидными связями, имеющие сложную
- 6. Классификация белков I. В зависимости от формы молекул белки подразделяются на фибриллярные и глобулярные. Молекулы фибриллярных
- 7. Классификация пептидов и белков. Глобулярные белки сложены в компактные глобулы. Водородные связи являются в основном внутримолекулярными,
- 8. Классификация пептидов и белков Строение белков определяет те функции, которые они выполняют в живых организмах. Фибриллярные
- 9. II. II. По составу белки делят на простые (неконъюгированные) и сложные (конъюгированные). По ряду характерных свойств
- 10. Простые белки: 1) Альбумины – растворимы в воде, не растворимы в конц. растворах солей. pl= 4,6-4,7.
- 11. коллагены - фибрилярные белки соединительной ткани. При длительном кипячении они растворяются в воде и при застудневании
- 12. Сложные белки : 1) Нуклеопротеины - содержат нуклеиновые кислоты. Наиболее изученными являются рибосомы, состоящие из нескольких
- 13. Гемопротеины - небелковый компонент этих протеидов - гем, построен из четырех пиррольных колец, с ними связан
- 14. 3) Металлопротеины - в состав простетической группы входят металлы. Хлорофилл - содержит гем, но вместо железа
- 15. Классификация пептидов и белков.
- 16. Функции белков Ферментативная функция Структурная функция. Питательная функция. Защитная функция. Транспортная функция. Регуляторная функция. Запасающая функция.
- 17. 2. Пространственное строение пептидов и белков
- 18. Среди большого числа гипотез о строении молекулы белка лишь одна выдержала испытание временем: полипептидная теория строения
- 19. Уровни структурной организации белка первичная структура – аминокислотная последовательность вторичная структура – локальные высокоупорядоченные конформации белковой
- 20. Пространственное строение пептидов и белков
- 21. Пространственное строение пептидов и белков Первичная структура пептидов и белков представляет собой полипептидную цепь последовательно связанных
- 22. Пептидная связь дипептид Пространственное строение пептидов и белков
- 23. Пространственное строение пептидов и белков
- 24. Пространственное строение пептидов и белков
- 25. Строение пептидной (амидной) группы Пространственное строение пептидов и белков В пептидной (амидной) группе –CONH атом углерода
- 26. С позиций электронного строения пептидная группа представляет собой трехцентровую, р,π -сопряженную систему, электронная плотность в которой
- 27. Наличие плоской сопряженной системы в пептидной группе является причиной затруднения вращения вокруг связи C-N, а углеродные
- 29. Изоэлектрическая точка пептидов и белков Как и аминокислоты, пептиды и белки являются амфотерными соединениями, содержащими и
- 30. Аминокислотная последовательность белков Первые исследования по выяснению аминокислотной последовательной белков были выполнены в Кембрижском университете Ф.Сенгером,
- 31. 20 аминокислот могут дать примерно 100000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000 белков, состоящих из 150 остатков аминокислот. Это астрономическое число –
- 32. Секвенирование биополимеров (белков и нуклеиновых кислот — ДНК и РНК) — определение их аминокислотной или нуклеотидной
- 33. Секвенирование ДНК Первичная структура любой белковой молекулы напрямую зависит от структуры ДНК-генома. Поэтому сначала выделяют ген,
- 34. Анализ первичной структуры пептидов Метод Эдмана Используя фенилизотиоцианат (Ph-NCS) последовательно отщепляют АК с N-конца и определяют
- 35. Фенилтиогидантоиновое производное N-концевой АК Пептид укороченный на 1 АК Пространственное строение пептидов и белков
- 36. Вторичная структура белка возникает за счет водородных связей между пептидными связями, что приводит к упорядоченному расположению
- 37. α-спираль Представляет собой полипептидную цепь расположенную в виде правозакрученной спирали. Внешне она похожа на слегка растянутую
- 38. Пространственное строение пептидов и белков
- 39. Водородные связи в α-спиралях -Вал-Фен-Тир-Ала-Лей- Первый Второй Третий Четвёртый Остатки АК: α-спираль – 413 спираль: остаток
- 40. α-спираль Правые α-спирали в полипептидной цепи стабилизируются водородными связями, где С=О группы связаны с лежащими от
- 41. β-слой Пространственное строение пептидов и белков
- 42. Пространственное строение пептидов и белков
- 43. Структура β-складчатых слоев водородные связи Тре-Ала-Гли-Сер-Асп Вал-Фен-Тир-Ала-Лей Пространственное строение пептидов и белков
- 44. Третичная структура белка возникает в результате взаимодействия между радикалами аминокислотных остатков полипептидных цепей. К таким взаимодействиям
- 45. Выделяют два общих типа третичной структуры: 1) В Фибриллярных белках (например, коллаген, эластин) молекулы которых имеют
- 46. Укладка α-спиралей и β-структуры с образованием глобулы Пространственное строение пептидов и белков
- 47. Взаимодействия остатков АК в третичной структуре АК в белковой глобуле взаимодействуют за счёт: Ковалентных связей (дисульфидные
- 48. Дисульфидные связи в цистине Пространственное строение пептидов и белков
- 49. Ионные связи Асп Лиз Пространственное строение пептидов и белков
- 50. Третичная структура ацилтрансфераза пиктатлиаза С Пространственное строение пептидов и белков
- 51. Третичная структура α и β цепи гемоглобина лошади Пространственное строение пептидов и белков
- 52. Домены Домены – глобулярные области в пределах одной белковой молекулы Домены соединены шарнирным участком Доменная структура
- 53. Четвертичная структура Четвертичная структура - способ укладки в пространстве отдельных полипептидных цепей, обладающих одинаковой первичной, вторичной,
- 54. Пространственное строение пептидов и белков 2130 одинаковых молекул белка расположены вокруг РНК вируса
- 55. 3. Физико-химические свойства пептидов и белков
- 56. Образуют коллоидные растворы. Большинство белков гидрофильны. Однако белковые молекулы имеют очень большие размеры, поэтому белки не
- 57. Несмотря на большую величину, многие белковые молекулы не осаждаются в водных растворах. Осаждению белковых молекул препятствуют
- 58. ФАКТОРЫ СТАБИЛИЗАЦИИ БЕЛКА В РАСТВОРЕ ГИДРАТНАЯ ОБОЛОЧКА - это слой молекул воды, определенным образом ориентированных на
- 59. Осаждение нативных белков 1. ВЫСАЛИВАНИЕ - это осаждение белков высокими концентрациями нейтральных солей щелочных и щелочноземельных
- 60. Белки менее гидрофильные, которые плохо удерживают воду гидратной оболочки, теряют ее раньше. Более гидрофильные белки требуют
- 61. 2. ПРИМЕНЕНИЕ ВОДООТНИМАЮЩИХ СРЕДСТВ. Такими средствами являются растворители, которые смешиваются с водой в любых соотношениях. Чаще
- 62. ДЕНАТУРАЦИЯ - это лишение белка eгo природных, нативных свойств, сопровождающееся разрушением четвертичной (если она была), третичной,
- 63. ФАКТОРЫ, ВЫЗЫВАЮЩИЕ ДЕНАТУРАЦИЮ БЕЛКОВ Физико-химические свойства пептидов и белков
- 64. ДЕЙСТВИЕ СОЛЕЙ ТЯЖЕЛЫХ МЕТАЛЛОВ. Образуют соединения с SН-группами белков. Ядовиты для человека и животных. В медицинской
- 65. Обратимость денатурации В пробирке (in vitгo) чаще всего это - необратимый процесс. Если же денатурированный белок
- 66. 4. Пептидный синтез
- 67. Чтобы соединить две аминокислоты пептидной связью, например получить метионилглицин, необходимо: 1) закрыть (защитить) аминогруппу метионина и
- 68. Пептидный синтез
- 69. Пептидный синтез
- 70. Пептидный синтез
- 71. Пептидный синтез
- 72. Пептидный синтез
- 73. Очень перспективный метод синтеза пептидных связей предложил в 1960 г. Меррифильд (США) метод твердофазного синтеза пептидов.
- 74. Пептидный синтез
- 75. 5. Методы выделения и очистки белка
- 76. Диализ Диализ используют для отделения низкомолекулярных примесей или замены состава среды. Метод основан на том, что
- 77. Гель-фильтрация Позволяет разделять белки по величине и форме молекул. Разделение проводят в хроматографических колонках, заполненных сферическими
- 78. Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия Является общепринятым методом определения гомогенности белковых препаратов. Метод
- 79. Под действием ДСН олигомерные белки диссоциируют на субъединицы и денатурируют. Развернутые полипептидные цепи связывают ДСН (примерно
- 81. Скачать презентацию

Цели лекции:
Обучающая - Сформировать знания о строении пептидов и белков и
Цели лекции:
Обучающая - Сформировать знания о строении пептидов и белков и

План
1. Классификация пептидов и белков. Основные функции белков.
2. Пространственное строение пептидов
План
1. Классификация пептидов и белков. Основные функции белков.
2. Пространственное строение пептидов

1. Классификация пептидов и белков
1. Классификация пептидов и белков

Белки и пептиды- полимерные азотсодержащие органические вещества состоящие из
Белки и пептиды- полимерные азотсодержащие органические вещества состоящие из

Классификация белков
I. В зависимости от формы молекул белки подразделяются на фибриллярные
Классификация белков
I. В зависимости от формы молекул белки подразделяются на фибриллярные

Классификация пептидов и белков.
Глобулярные белки сложены в компактные глобулы. Водородные
Классификация пептидов и белков.
Глобулярные белки сложены в компактные глобулы. Водородные

Классификация пептидов и белков
Строение белков определяет те функции, которые они
Классификация пептидов и белков
Строение белков определяет те функции, которые они

II. II. По составу белки делят на простые (неконъюгированные) и сложные
II. II. По составу белки делят на простые (неконъюгированные) и сложные

Простые белки:
1) Альбумины – растворимы в воде, не
Простые белки:
1) Альбумины – растворимы в воде, не

коллагены - фибрилярные белки соединительной ткани. При длительном кипячении они
коллагены - фибрилярные белки соединительной ткани. При длительном кипячении они

Сложные белки :
1) Нуклеопротеины - содержат нуклеиновые кислоты. Наиболее изученными
Сложные белки :
1) Нуклеопротеины - содержат нуклеиновые кислоты. Наиболее изученными

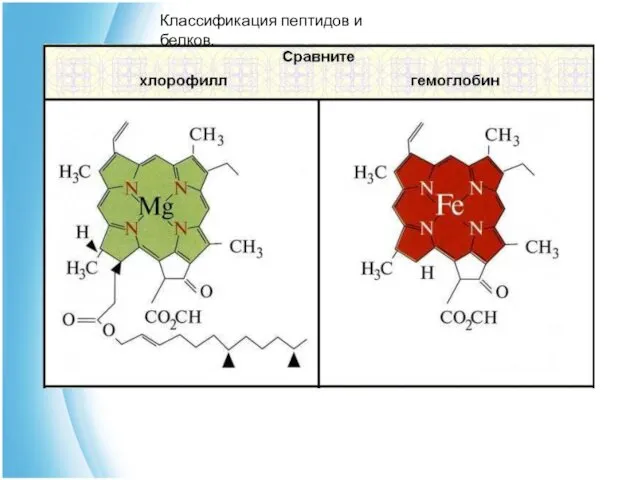
Гемопротеины - небелковый компонент этих протеидов - гем, построен из
Гемопротеины - небелковый компонент этих протеидов - гем, построен из

3) Металлопротеины - в состав простетической группы входят металлы.
Хлорофилл - содержит
3) Металлопротеины - в состав простетической группы входят металлы.
Хлорофилл - содержит
Классификация пептидов и белков.
Классификация пептидов и белков.

Функции белков
Ферментативная функция
Структурная функция.
Питательная функция.
Защитная
Функции белков
Ферментативная функция
Структурная функция.
Питательная функция.
Защитная

2. Пространственное строение пептидов и белков
2. Пространственное строение пептидов и белков

Среди большого числа гипотез о строении молекулы белка лишь одна выдержала
Среди большого числа гипотез о строении молекулы белка лишь одна выдержала

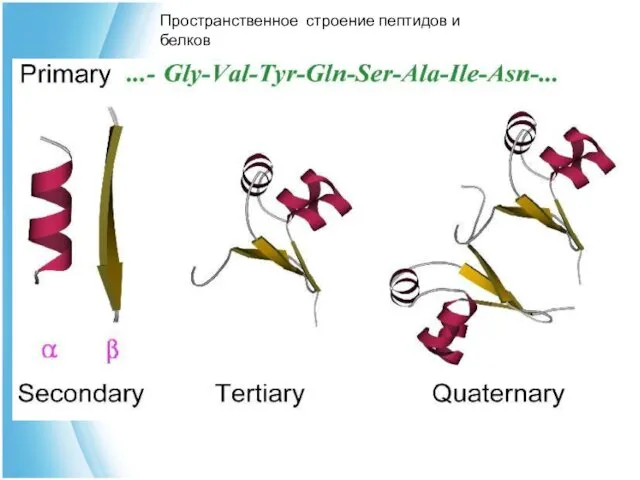
Уровни структурной организации белка
первичная структура –
аминокислотная последовательность
вторичная структура – локальные
Уровни структурной организации белка
первичная структура –
аминокислотная последовательность
вторичная структура – локальные
Пространственное строение пептидов и белков
Пространственное строение пептидов и белков
Пространственное строение пептидов и белков
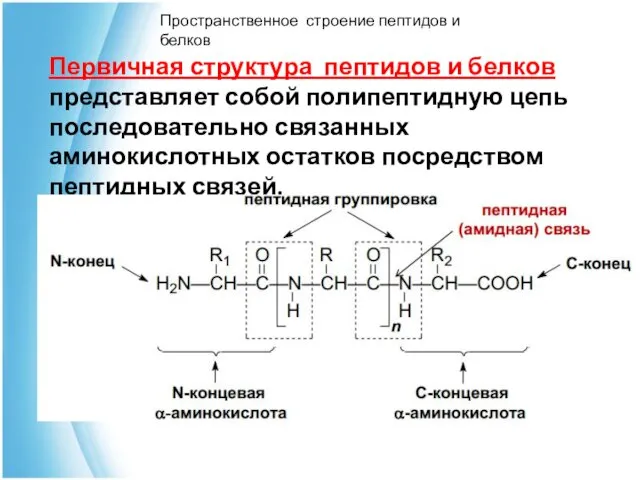
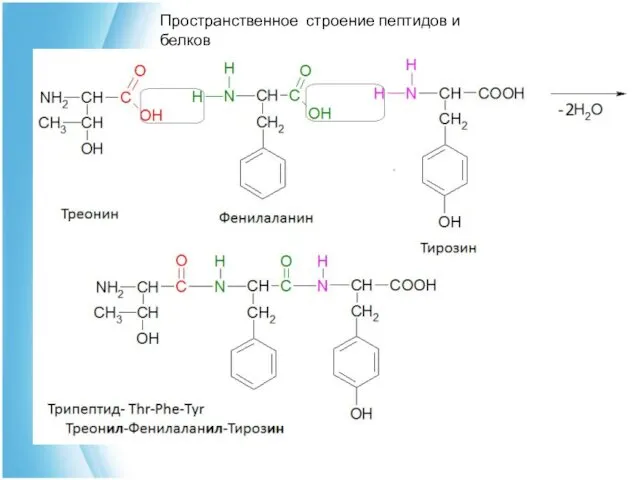
Первичная структура пептидов и белков представляет собой
Пространственное строение пептидов и белков
Первичная структура пептидов и белков представляет собой
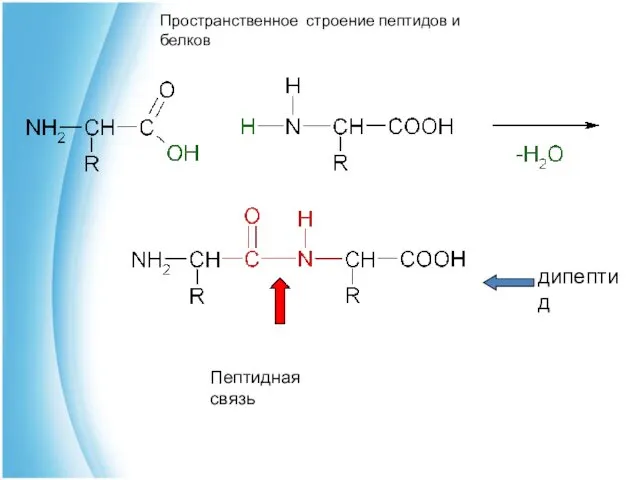
Пептидная связь
дипептид
Пространственное строение пептидов и белков
Пептидная связь
дипептид
Пространственное строение пептидов и белков
Пространственное строение пептидов и белков
Пространственное строение пептидов и белков
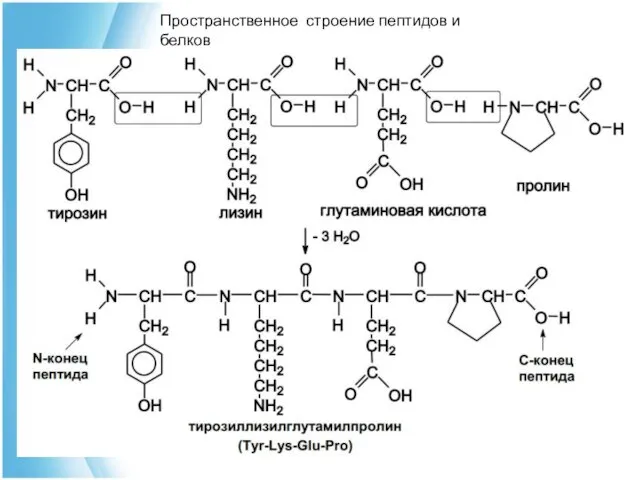
Пространственное строение пептидов и белков
Пространственное строение пептидов и белков
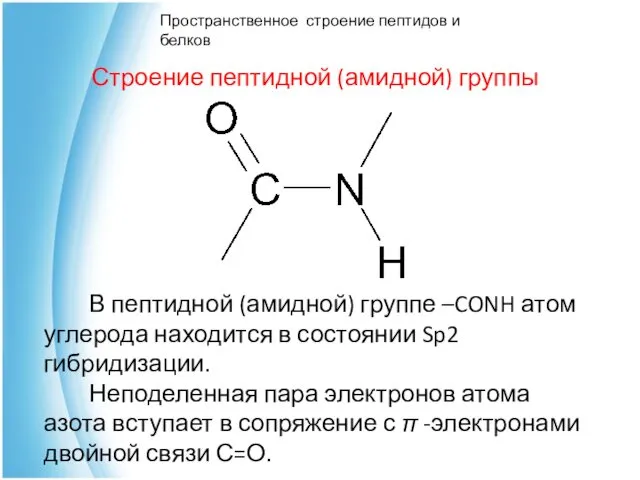
Строение пептидной (амидной) группы
Пространственное строение пептидов и белков
В пептидной (амидной)
Строение пептидной (амидной) группы
Пространственное строение пептидов и белков
В пептидной (амидной)
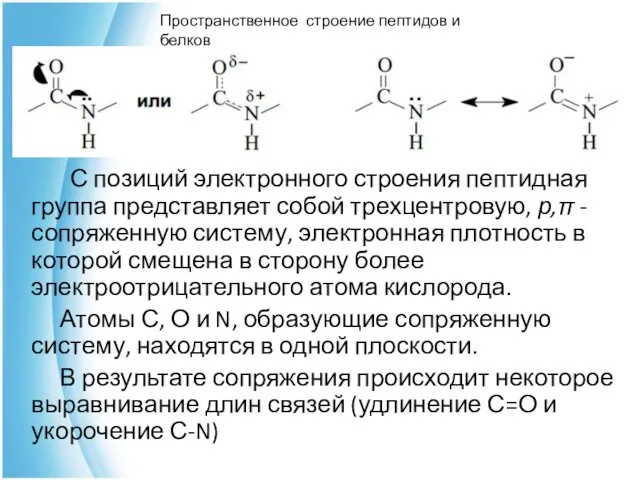
С позиций электронного строения пептидная группа представляет собой трехцентровую, р,π
С позиций электронного строения пептидная группа представляет собой трехцентровую, р,π
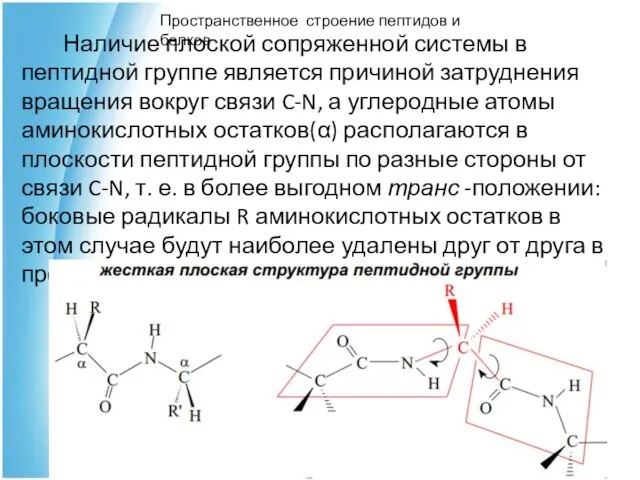
Наличие плоской сопряженной системы в пептидной группе является причиной затруднения
Наличие плоской сопряженной системы в пептидной группе является причиной затруднения
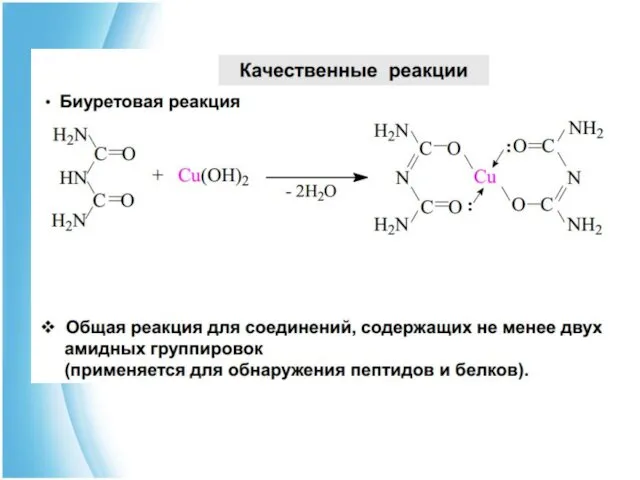

Изоэлектрическая точка пептидов и белков
Как и аминокислоты, пептиды и
Изоэлектрическая точка пептидов и белков
Как и аминокислоты, пептиды и

Аминокислотная последовательность белков
Первые исследования по выяснению аминокислотной последовательной белков
Аминокислотная последовательность белков
Первые исследования по выяснению аминокислотной последовательной белков

20 аминокислот могут дать примерно 100000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000 белков, состоящих из 150
20 аминокислот могут дать примерно 100000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000 белков, состоящих из 150

Секвенирование биополимеров (белков и нуклеиновых кислот — ДНК и РНК) — определение их аминокислотной или нуклеотидной последовательности (от лат. sequentum — последовательность). В результате
Секвенирование биополимеров (белков и нуклеиновых кислот — ДНК и РНК) — определение их аминокислотной или нуклеотидной последовательности (от лат. sequentum — последовательность). В результате

Секвенирование ДНК
Первичная структура любой белковой молекулы напрямую зависит
Секвенирование ДНК
Первичная структура любой белковой молекулы напрямую зависит

Анализ первичной структуры пептидов
Метод Эдмана
Используя фенилизотиоцианат (Ph-NCS) последовательно отщепляют АК
Анализ первичной структуры пептидов
Метод Эдмана
Используя фенилизотиоцианат (Ph-NCS) последовательно отщепляют АК
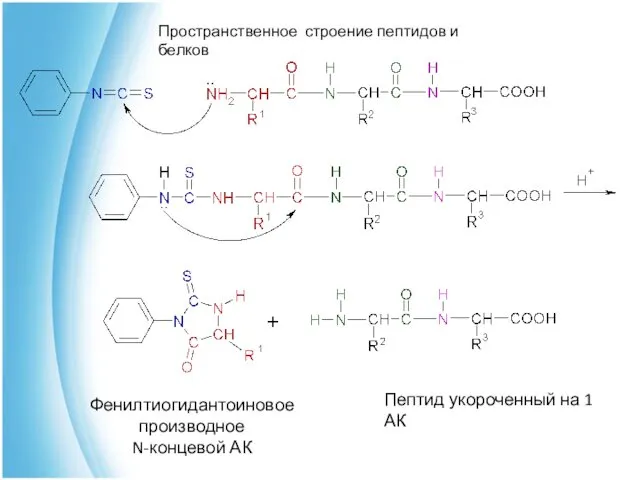
Фенилтиогидантоиновое производное
N-концевой АК
Пептид укороченный на 1 АК
Пространственное строение пептидов и
Фенилтиогидантоиновое производное
N-концевой АК
Пептид укороченный на 1 АК
Пространственное строение пептидов и

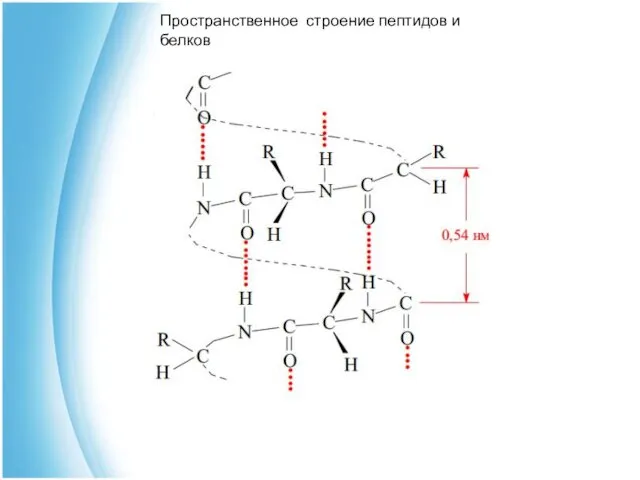
Вторичная структура белка возникает за счет водородных связей между пептидными
Вторичная структура белка возникает за счет водородных связей между пептидными

α-спираль
Представляет собой полипептидную цепь расположенную в виде правозакрученной спирали.
α-спираль
Представляет собой полипептидную цепь расположенную в виде правозакрученной спирали.
Пространственное строение пептидов и белков
Пространственное строение пептидов и белков
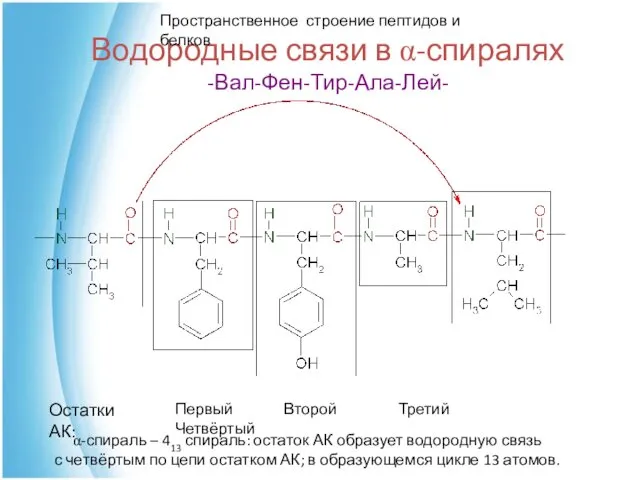
Водородные связи в α-спиралях
-Вал-Фен-Тир-Ала-Лей-
Первый Второй Третий Четвёртый
Остатки АК:
α-спираль – 413 спираль:
Водородные связи в α-спиралях
-Вал-Фен-Тир-Ала-Лей-
Первый Второй Третий Четвёртый
Остатки АК:
α-спираль – 413 спираль:
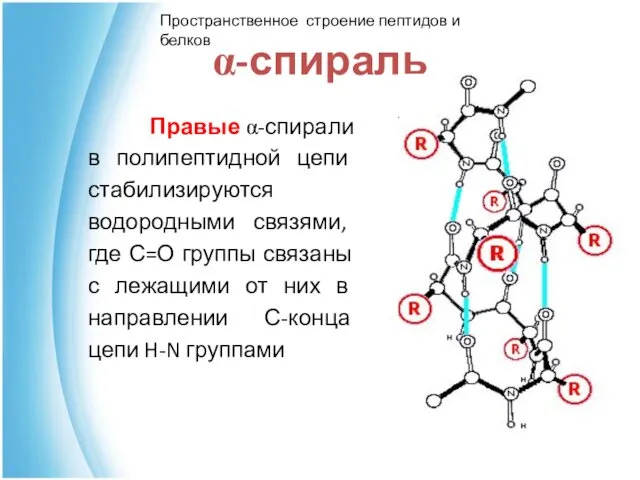
α-спираль
Правые α-спирали в полипептидной цепи стабилизируются водородными связями, где С=О
α-спираль
Правые α-спирали в полипептидной цепи стабилизируются водородными связями, где С=О
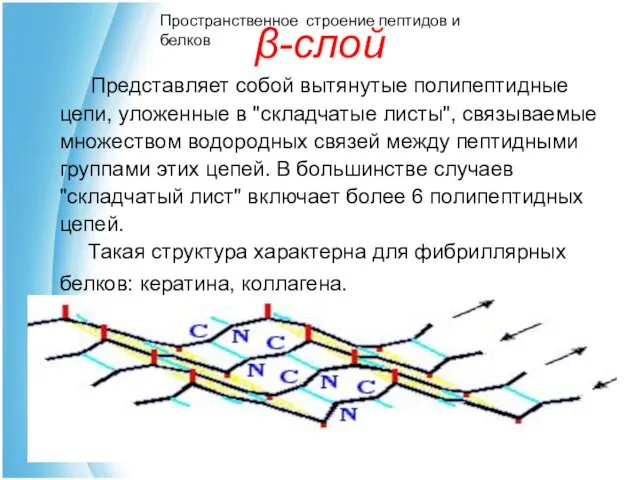
β-слой
Пространственное строение пептидов и белков
β-слой
Пространственное строение пептидов и белков
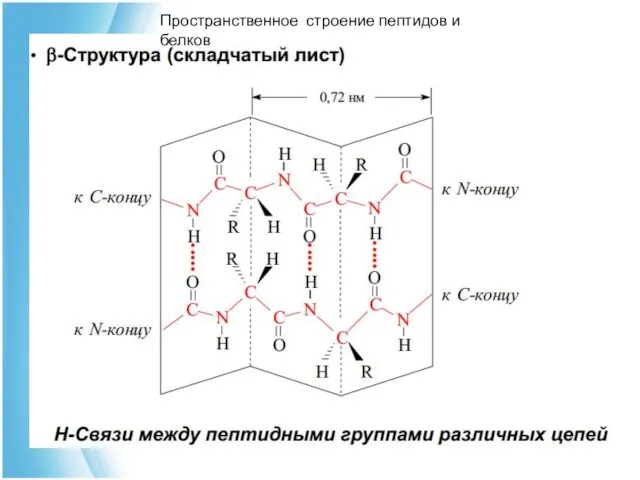
Пространственное строение пептидов и белков
Пространственное строение пептидов и белков
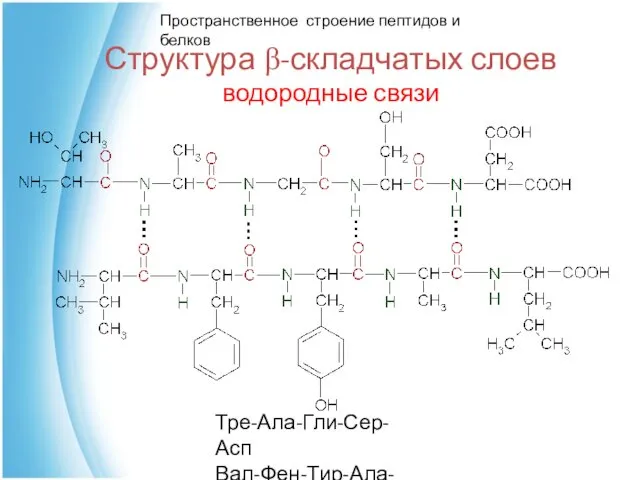
Структура β-складчатых слоев
водородные связи
Тре-Ала-Гли-Сер-Асп
Вал-Фен-Тир-Ала-Лей
Пространственное строение пептидов и белков
Структура β-складчатых слоев
водородные связи
Тре-Ала-Гли-Сер-Асп
Вал-Фен-Тир-Ала-Лей
Пространственное строение пептидов и белков

Третичная структура белка возникает в результате взаимодействия между радикалами аминокислотных
Третичная структура белка возникает в результате взаимодействия между радикалами аминокислотных

Выделяют два общих типа третичной структуры:
1) В Фибриллярных белках (например,
Выделяют два общих типа третичной структуры:
1) В Фибриллярных белках (например,
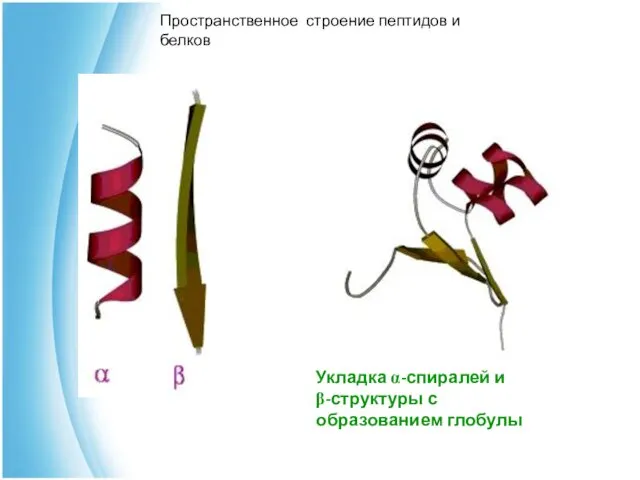
Укладка α-спиралей и
β-структуры с образованием глобулы
Пространственное строение пептидов и белков
Укладка α-спиралей и
β-структуры с образованием глобулы
Пространственное строение пептидов и белков

Взаимодействия остатков АК в третичной структуре
АК в белковой глобуле взаимодействуют за
Взаимодействия остатков АК в третичной структуре
АК в белковой глобуле взаимодействуют за
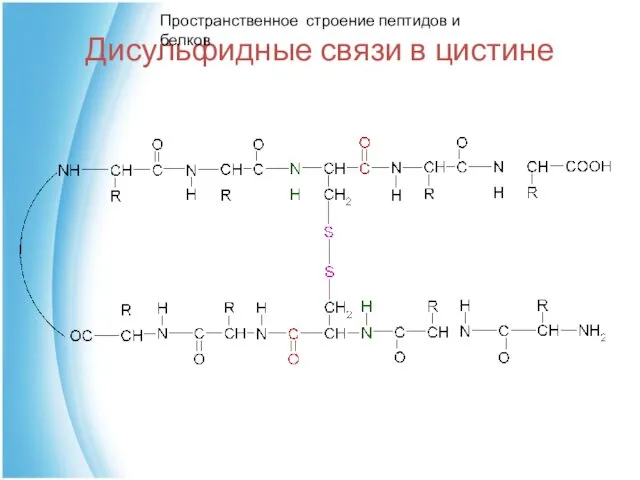
Дисульфидные связи в цистине
Пространственное строение пептидов и белков
Дисульфидные связи в цистине
Пространственное строение пептидов и белков
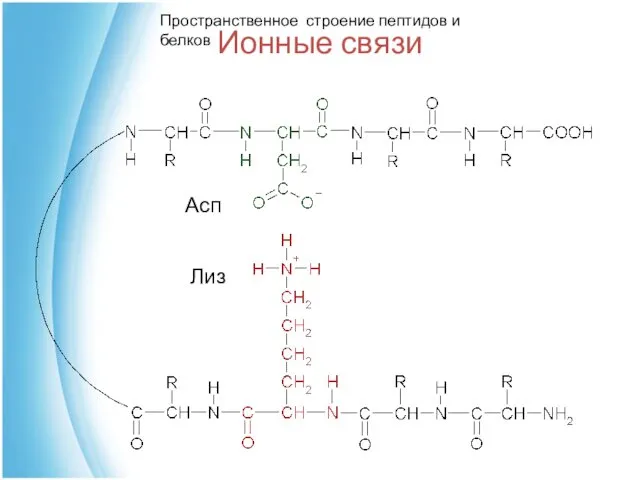
Ионные связи
Асп
Лиз
Пространственное строение пептидов и белков
Ионные связи
Асп
Лиз
Пространственное строение пептидов и белков
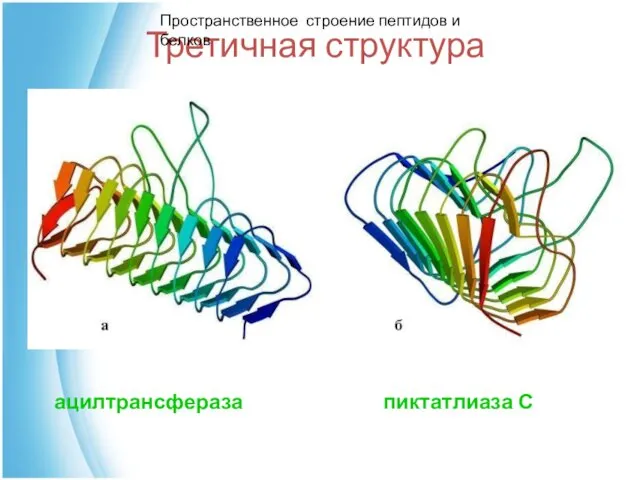
Третичная структура
ацилтрансфераза пиктатлиаза С
Пространственное строение пептидов и белков
Третичная структура
ацилтрансфераза пиктатлиаза С
Пространственное строение пептидов и белков
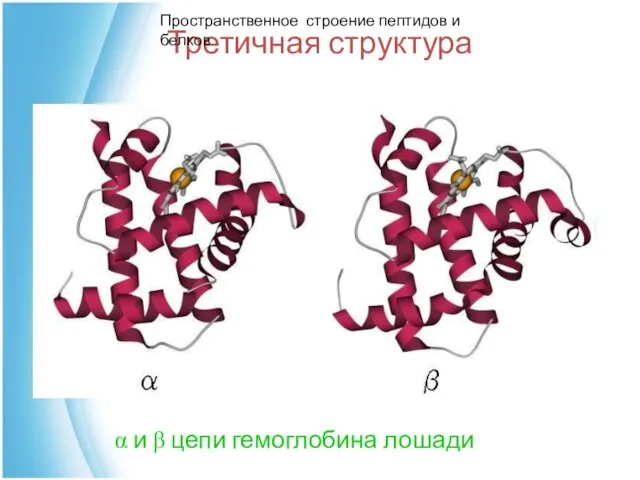
Третичная структура
α и β цепи гемоглобина лошади
Пространственное строение пептидов и белков
Третичная структура
α и β цепи гемоглобина лошади
Пространственное строение пептидов и белков

Домены
Домены – глобулярные области в пределах одной белковой молекулы
Домены соединены
Домены
Домены – глобулярные области в пределах одной белковой молекулы
Домены соединены

Четвертичная структура
Четвертичная структура - способ укладки в пространстве отдельных
Четвертичная структура
Четвертичная структура - способ укладки в пространстве отдельных

Пространственное строение пептидов и белков
2130 одинаковых молекул белка расположены вокруг РНК
Пространственное строение пептидов и белков
2130 одинаковых молекул белка расположены вокруг РНК

3. Физико-химические свойства пептидов и белков
3. Физико-химические свойства пептидов и белков

Образуют коллоидные растворы.
Большинство белков гидрофильны. Однако белковые молекулы
Образуют коллоидные растворы.
Большинство белков гидрофильны. Однако белковые молекулы

Несмотря на большую величину, многие белковые молекулы не осаждаются в
Несмотря на большую величину, многие белковые молекулы не осаждаются в

ФАКТОРЫ СТАБИЛИЗАЦИИ БЕЛКА В РАСТВОРЕ
ГИДРАТНАЯ ОБОЛОЧКА - это слой
ФАКТОРЫ СТАБИЛИЗАЦИИ БЕЛКА В РАСТВОРЕ
ГИДРАТНАЯ ОБОЛОЧКА - это слой

Осаждение нативных белков
1. ВЫСАЛИВАНИЕ - это осаждение белков высокими
Осаждение нативных белков
1. ВЫСАЛИВАНИЕ - это осаждение белков высокими

Белки менее гидрофильные, которые плохо удерживают воду гидратной оболочки, теряют
Белки менее гидрофильные, которые плохо удерживают воду гидратной оболочки, теряют

2. ПРИМЕНЕНИЕ ВОДООТНИМАЮЩИХ СРЕДСТВ.
Такими средствами являются растворители, которые смешиваются
2. ПРИМЕНЕНИЕ ВОДООТНИМАЮЩИХ СРЕДСТВ.
Такими средствами являются растворители, которые смешиваются

ДЕНАТУРАЦИЯ - это лишение белка eгo природных, нативных свойств, сопровождающееся
ДЕНАТУРАЦИЯ - это лишение белка eгo природных, нативных свойств, сопровождающееся

ФАКТОРЫ, ВЫЗЫВАЮЩИЕ ДЕНАТУРАЦИЮ БЕЛКОВ
Физико-химические свойства пептидов и белков
ФАКТОРЫ, ВЫЗЫВАЮЩИЕ ДЕНАТУРАЦИЮ БЕЛКОВ
Физико-химические свойства пептидов и белков

ДЕЙСТВИЕ СОЛЕЙ ТЯЖЕЛЫХ МЕТАЛЛОВ. Образуют соединения с SН-группами белков. Ядовиты для
ДЕЙСТВИЕ СОЛЕЙ ТЯЖЕЛЫХ МЕТАЛЛОВ. Образуют соединения с SН-группами белков. Ядовиты для

Обратимость денатурации
В пробирке (in vitгo) чаще всего это -
Обратимость денатурации
В пробирке (in vitгo) чаще всего это -

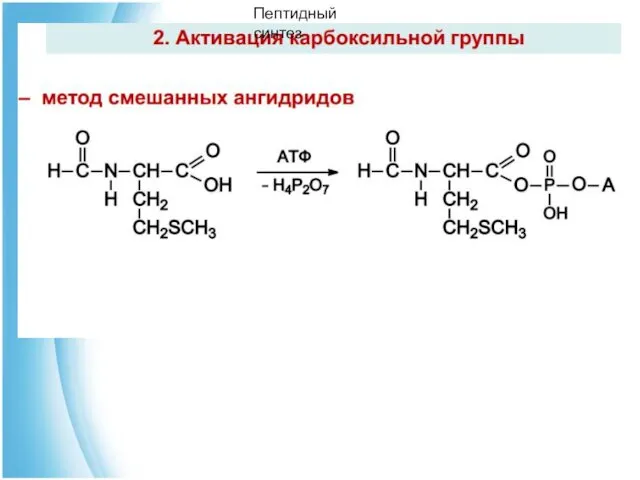
4. Пептидный синтез
4. Пептидный синтез

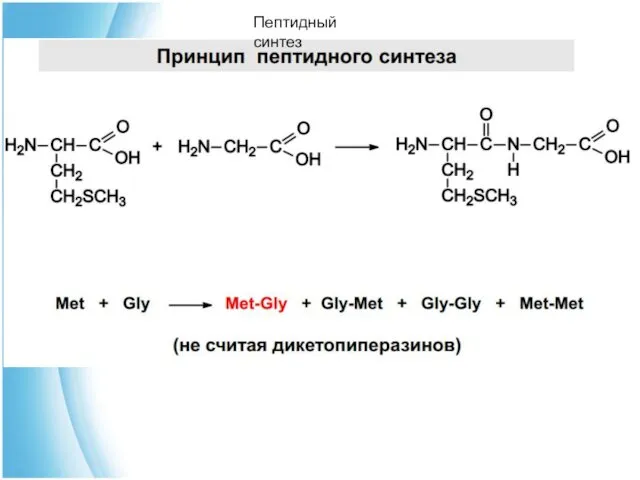
Чтобы соединить две аминокислоты пептидной связью, например получить метионилглицин, необходимо:
Чтобы соединить две аминокислоты пептидной связью, например получить метионилглицин, необходимо:
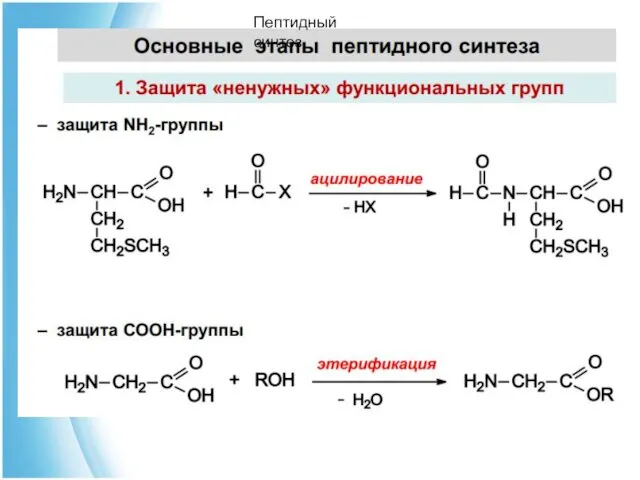
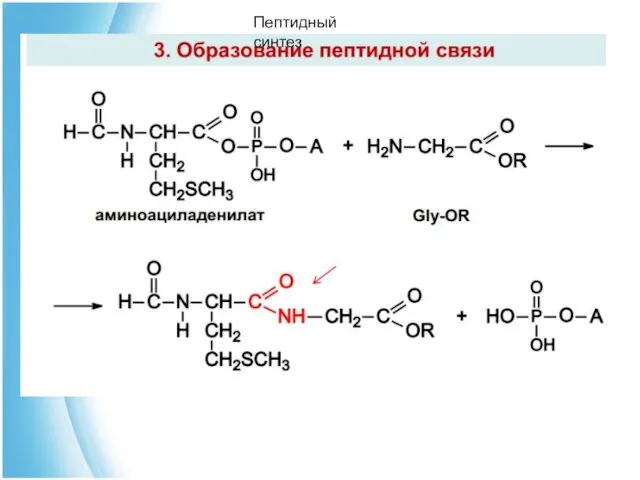
Пептидный синтез
Пептидный синтез
Пептидный синтез
Пептидный синтез
Пептидный синтез
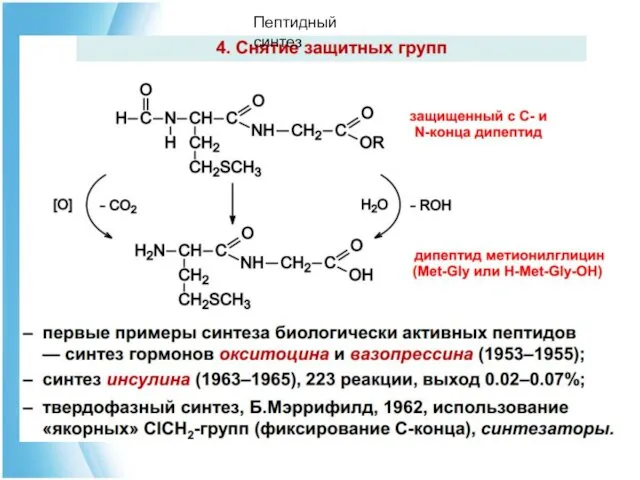
Пептидный синтез
Пептидный синтез
Пептидный синтез
Пептидный синтез
Пептидный синтез

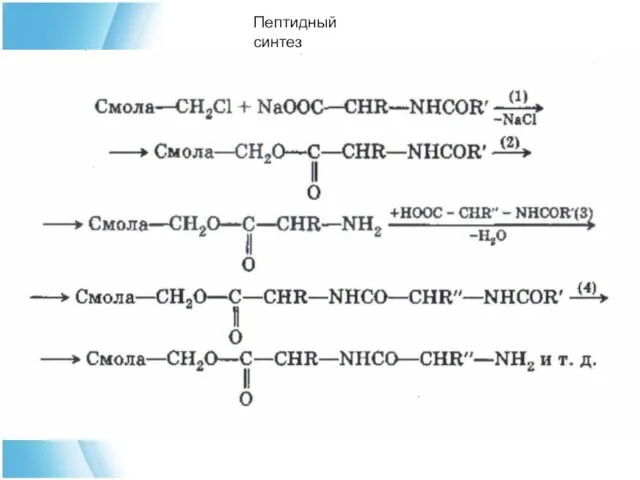
Очень перспективный метод синтеза пептидных связей предложил в 1960 г. Меррифильд
Очень перспективный метод синтеза пептидных связей предложил в 1960 г. Меррифильд
Пептидный синтез
Пептидный синтез

5. Методы выделения и очистки белка
5. Методы выделения и очистки белка
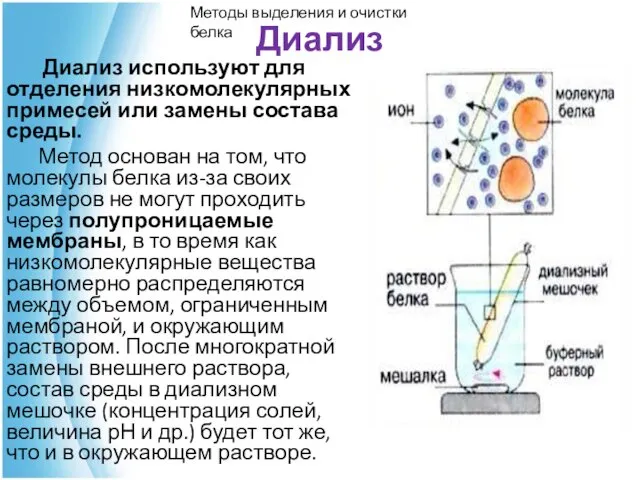
Диализ
Диализ используют для отделения низкомолекулярных примесей или замены состава
Диализ
Диализ используют для отделения низкомолекулярных примесей или замены состава
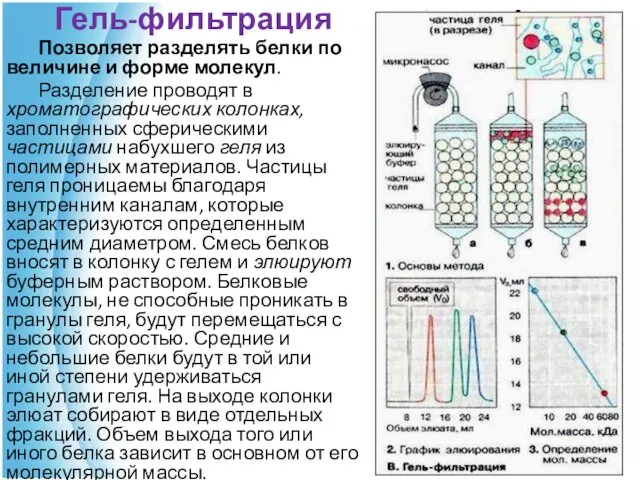
Гель-фильтрация
Позволяет разделять белки по величине и форме молекул.
Разделение проводят
Гель-фильтрация
Позволяет разделять белки по величине и форме молекул.
Разделение проводят
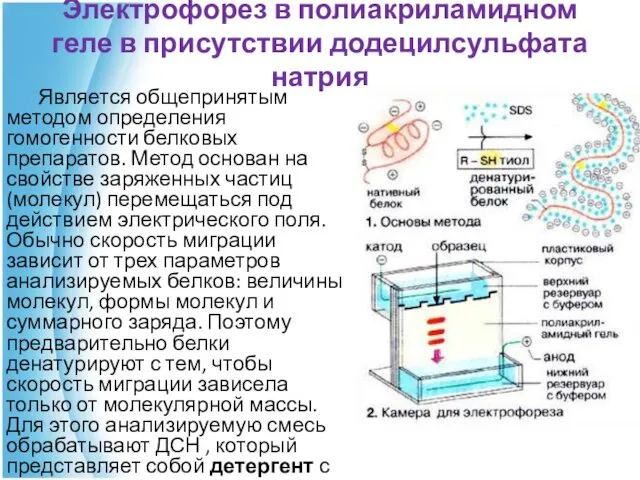
Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия
Является общепринятым методом
Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия
Является общепринятым методом
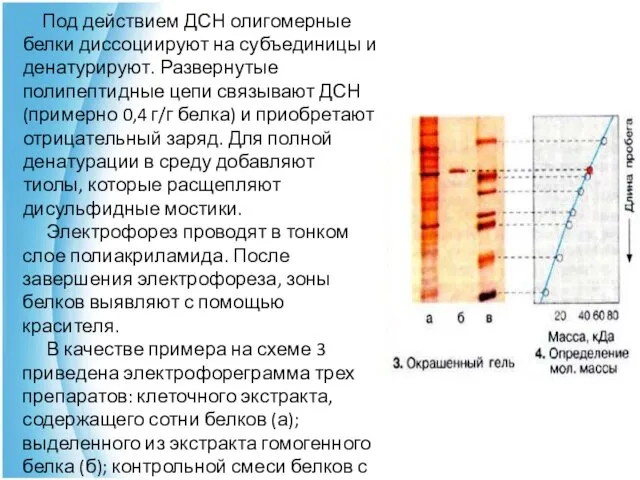
Под действием ДСН олигомерные белки диссоциируют на субъединицы и денатурируют.
Под действием ДСН олигомерные белки диссоциируют на субъединицы и денатурируют.
 Номенклатура органических соединений
Номенклатура органических соединений Группа галогенов в периодической системе
Группа галогенов в периодической системе Неорганическая химия
Неорганическая химия Лекарственные препараты гетероциклического строения, производные фурана, пиррола, тиазола, бензтиазола, тиадиазола. (Тема 1)
Лекарственные препараты гетероциклического строения, производные фурана, пиррола, тиазола, бензтиазола, тиадиазола. (Тема 1) Гидролиз
Гидролиз Углеводы. Классификация, общая характеристика
Углеводы. Классификация, общая характеристика Атомно-кристаллическое строение материалов
Атомно-кристаллическое строение материалов Бионеорганическая химия. Жидкие среды организма. (Лекция 2)
Бионеорганическая химия. Жидкие среды организма. (Лекция 2) Химия и стирка
Химия и стирка  Термодинамика. Химическая термодинамика
Термодинамика. Химическая термодинамика Цикл три і дикарбонових кислот (Цикл Кребса)
Цикл три і дикарбонових кислот (Цикл Кребса) Physical chemistry of nanostructured systems.( lecture no. 7)
Physical chemistry of nanostructured systems.( lecture no. 7) Золь-гель метод получения газочувствительных слоев диоксида олова
Золь-гель метод получения газочувствительных слоев диоксида олова Effect receiving modes, and heat treatment on the structure and properties of the magnetic powder of strontium hexaferrite
Effect receiving modes, and heat treatment on the structure and properties of the magnetic powder of strontium hexaferrite Жири
Жири  Вычисление массовой доли растворенного вещества в растворе. Вычисление количества вещества, массы или объема вещества
Вычисление массовой доли растворенного вещества в растворе. Вычисление количества вещества, массы или объема вещества Крекинг нефтепродуктов
Крекинг нефтепродуктов Сера. Сера в природе. Применение серы. Сероводород и сульфиды
Сера. Сера в природе. Применение серы. Сероводород и сульфиды Геохимические классификации элементов
Геохимические классификации элементов Неметаллы. Смотр знаний. Для учащихся 9 классов
Неметаллы. Смотр знаний. Для учащихся 9 классов Карбонильные соединения – альдегиды
Карбонильные соединения – альдегиды ПРИРОДОВЕДЕНИЕ 5 КЛАСС
ПРИРОДОВЕДЕНИЕ 5 КЛАСС Карбонильные соединения
Карбонильные соединения Функциональные производные карбоновых кислот
Функциональные производные карбоновых кислот Критерии оценки качества природного газа
Критерии оценки качества природного газа Строение атома алюминия. Физические и химические свойства алюминия
Строение атома алюминия. Физические и химические свойства алюминия Дегтярева М.О. Московская область г. Королёв АОУ ЛНИП 242-645-771
Дегтярева М.О. Московская область г. Королёв АОУ ЛНИП 242-645-771 d-элементы
d-элементы