- Глобальная регуляция экспрессии генов

Содержание
- 2. GR1.1 Локализация эу-(светлые части хромосом) и гетерохроматина (интенсивно окрашенные участки) в кариотипе дрозофилы по результатам С-окрашивания.
- 3. 15 УПАКОВКА ДНК В ХРОМОСОМАХ. ГИСТОНЫ Типичные характеристики гистонов млекопитающих ТИП Число Мм, кДа Число Lys/Arg
- 4. 16 УПАКОВКА ДНК В ХРОМОСОМАХ «бусы на нитке» хроматиновые фиблиллы вытянутые петли компактные петли
- 5. 16.1 УПАКОВКА ДНК В ХРОМОСОМАХ Электронная микрофотография одиночной хроматиды митотитеской хромосомы насекомого. Специальная обработка позволяет визуализировать
- 6. GR1.2 Метилирование ДНК Активность транскрипции зависит от степени метилирования цитозина в динуклеотидных парах оснований: 5’-CG-3’ 3’-GC-3’
- 7. GR2 В тканях эмбриона до его имплантации обе хромосомы активны и не метилированы. После имплантации экспрессия
- 8. СУДЬБА СИНТЕЗИРОВАННЫХ БЕЛКОВ 105 Посттрансляционная модификация Превращение препроинсулина в инсулин сигнальный пептид A B C препроинсулин
- 9. 106 Доставка эукариотических белков к клеточным мембранам и проникновение через них Эндоплазматический ретикулум (ЭР) - непрерывная
- 10. 107 Котрансляционный транспорт полипептидных цепей сигнальная последовательность
- 11. 108 Направление синтезируемых полипептидных цепей в просвет ЭР Транспорт мембранных и секретируемых белков в просвет ЭР
- 12. Scheme of the eubacterial complex Structure of the E. coli signal recognition particle bound to a
- 13. 109 Характерные N-концевые сигнальные последовательности эукариотических и прокариотических белков Длина сигнальной последовательности составляет от 15 до
- 14. 110 Транспорт белков от ЭР к аппарату Голъджи и из него Белки направляются к лизосомам, плазматическим
- 15. 111 Транспорт белков от ЭР к аппарату Гольджи и из него модификация транспорт
- 16. 112 Транспорт белков в эукариотические клеточные органеллы
- 17. Mechanisms of Nuclear Import and Export Gene Expression in Prokaryotes: DNA ? RNA ? Protein Gene
- 18. 112.5 Транспорт белков в ядро (nuclear import) Proteins to be imported into the nucleus have Nuclear
- 19. 112.6 Транспорт из ядра (nuclear export)
- 20. The classical nuclear import cycle. Allison Lange et al. J. Biol. Chem. 2007;282:5101-5105 ©2007 by American
- 21. 113 Клеточная стенка грамположительных (А) и грамотрицательных (Б) эубактерий: 1 — цитоплазматическая мембрана; 2 — пептидогликан;
- 22. 113.1 Строение клеточных стенок
- 23. 113.2 Строение клеточной стенки грам-отрицательных бактерий
- 24. 113.3 Строение клеточной стенки грам-положительных бактерий
- 25. 114 Характерные N-концевые сигнальные последовательности эукариотических и прокариотических белков Длина сигнальной последовательности составляет от 15 до
- 26. СТРУКТУРА ГЕНОМОВ
- 27. Геном - суммарная ДНК одного набора хромосом и внехромосомных генетических элементов организма. 114.5
- 28. 115 СТРУКТУРНЫЕ ОСОБЕННОСТИ ПРОКАРИОТИЧЕСКИХ И ЭУКАРИОТИЧЕСКИХ ГЕНОВ Сегменты ДНК, которые транскрибируются с образованием одной молекулы РНК
- 29. 116 ЭУКАРИОТИЧЕСКИЕ ГЕНЫ Гены класса I кодируют 5,8S-, 18S- и 28S-pPHK, транскрибируются РНК-полимеразой I (Pol I).
- 30. 117 Регуляторные участки гена транскрибируемого Pol II Фактор транскрипции - любой белок необходимый для инициации транскрипции,
- 31. 118 Расположение белков в типичном промоторе эукариот Общие факторы транскрипции PolII к настоящему времени выделены и
- 32. 119 Специфические факторы транскрипции Специфические факторы транскрипции должны обладать двумя важнейшими свойствами: 1) опознавать специфические последовательности-мишени,
- 33. 120 Специфические мотивы связывания с ДНК «Цинковые пальцы» - zinc finger domain (a) Организация домена эстрогенового
- 34. 121 Специфические мотивы связывания с ДНК Спираль-поворот-спираль - helix-turn-helix Взаимодействие димера репрессора λ с ДНК. Каждый
- 35. 122 Специфические мотивы связывания с ДНК Структура AP-1/DNA комплекса. AP-1 – димер, сформированный транскрипционным фактором Jun
- 36. 122.1 Специфические мотивы связывания с ДНК Фактор транскрипции MyoD. Структура мотива «helix-loop-helix». PDB ID: 1MDY Спираль-петля-спираль
- 37. 123 Энхансеры Энхансерные последовательности служат в качестве специфических участков связывания особых регуляторных белков, активирующих процесс транскрипции
- 38. Инсуляторы Только в пределах участка между двумя инсуляторами энхансерные последовательности, связавшись с белками-активаторами, могут образовать петлю
- 39. 125 Терминация транскрипции Механизмы, которые определяют 3'-конец соответствующих функциональных РНК-продуктов, уникальны для каждой полимеразной системы Сигналом
- 40. 126 Транскрипция Pol II многих генов белков эукариот осуществляется во множественных сайтах последовательности ДНК протяженностью в
- 41. 127 Экзоны и интроны Экзон-интронное строение некоторых генов человека Распределение частот встречаемости генов с различным числом
- 43. Скачать презентацию
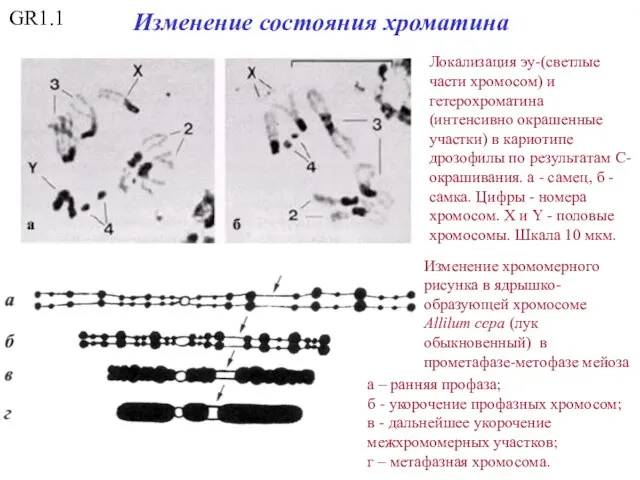
GR1.1
Локализация эу-(светлые части хромосом) и гетерохроматина (интенсивно окрашенные участки) в кариотипе
GR1.1
Локализация эу-(светлые части хромосом) и гетерохроматина (интенсивно окрашенные участки) в кариотипе
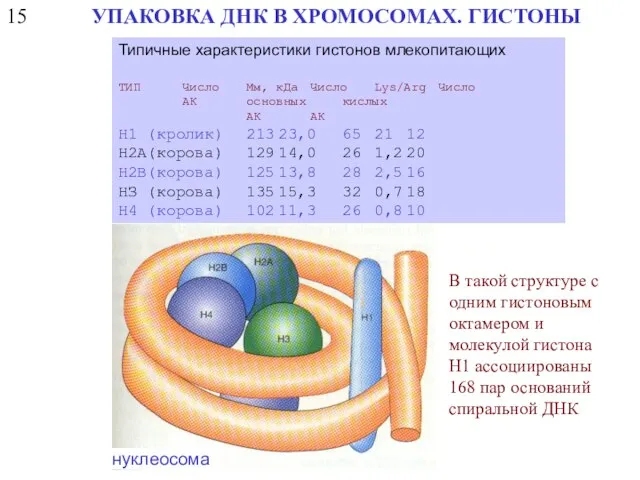
15
УПАКОВКА ДНК В ХРОМОСОМАХ. ГИСТОНЫ
Типичные характеристики гистонов млекопитающих
ТИП Число Мм, кДа Число
15
УПАКОВКА ДНК В ХРОМОСОМАХ. ГИСТОНЫ
Типичные характеристики гистонов млекопитающих
ТИП Число Мм, кДа Число
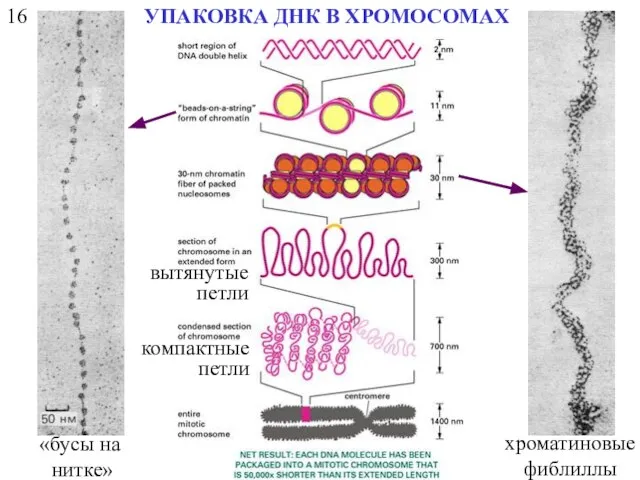
16
УПАКОВКА ДНК В ХРОМОСОМАХ
«бусы на
нитке»
хроматиновые фиблиллы
вытянутые петли
компактные петли
16
УПАКОВКА ДНК В ХРОМОСОМАХ
«бусы на
нитке»
хроматиновые фиблиллы
вытянутые петли
компактные петли
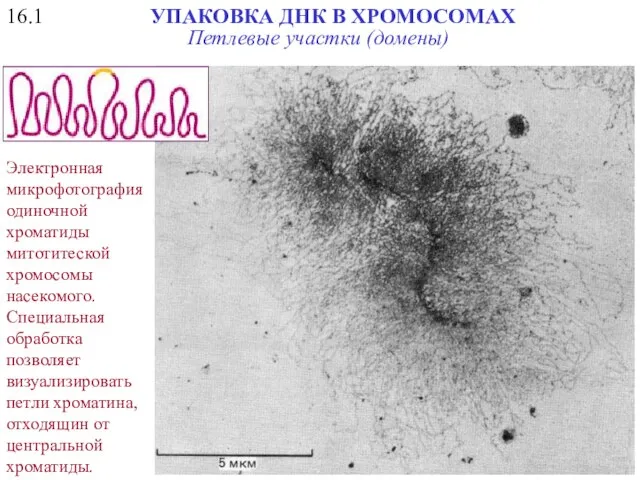
16.1
УПАКОВКА ДНК В ХРОМОСОМАХ
Электронная микрофотография одиночной хроматиды митотитеской хромосомы насекомого. Специальная
16.1
УПАКОВКА ДНК В ХРОМОСОМАХ
Электронная микрофотография одиночной хроматиды митотитеской хромосомы насекомого. Специальная

GR1.2
Метилирование ДНК
Активность транскрипции зависит от степени метилирования цитозина в динуклеотидных парах
GR1.2
Метилирование ДНК
Активность транскрипции зависит от степени метилирования цитозина в динуклеотидных парах
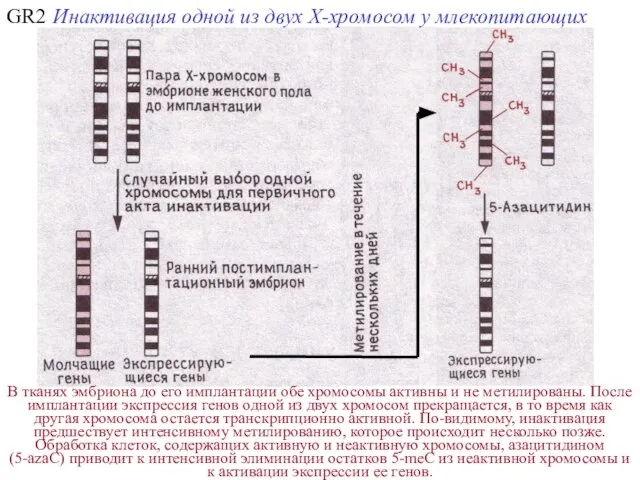
GR2
В тканях эмбриона до его имплантации обе хромосомы активны и не
GR2
В тканях эмбриона до его имплантации обе хромосомы активны и не
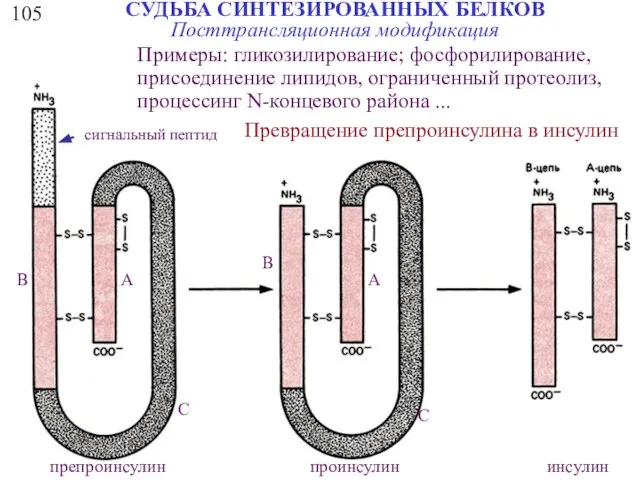
СУДЬБА СИНТЕЗИРОВАННЫХ БЕЛКОВ
105
Посттрансляционная модификация
Превращение препроинсулина в инсулин
сигнальный пептид
A
B
C
препроинсулин
проинсулин
инсулин
C
A
B
Примеры: гликозилирование; фосфорилирование,
СУДЬБА СИНТЕЗИРОВАННЫХ БЕЛКОВ
105
Посттрансляционная модификация
Превращение препроинсулина в инсулин
сигнальный пептид
A
B
C
препроинсулин
проинсулин
инсулин
C
A
B
Примеры: гликозилирование; фосфорилирование,
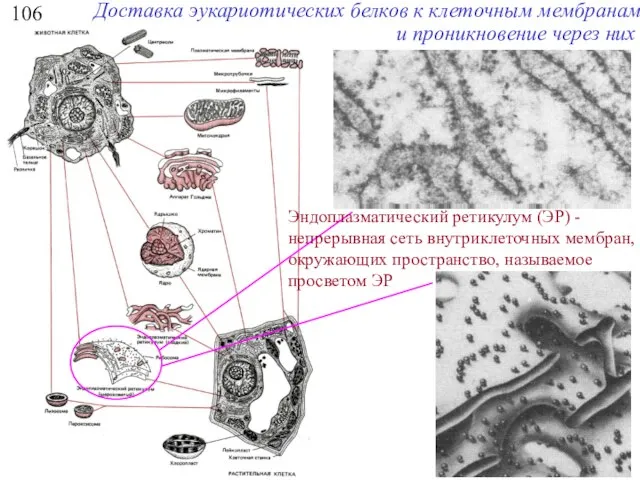
106
Доставка эукариотических белков к клеточным мембранам и проникновение через них
Эндоплазматический ретикулум
106
Доставка эукариотических белков к клеточным мембранам и проникновение через них
Эндоплазматический ретикулум
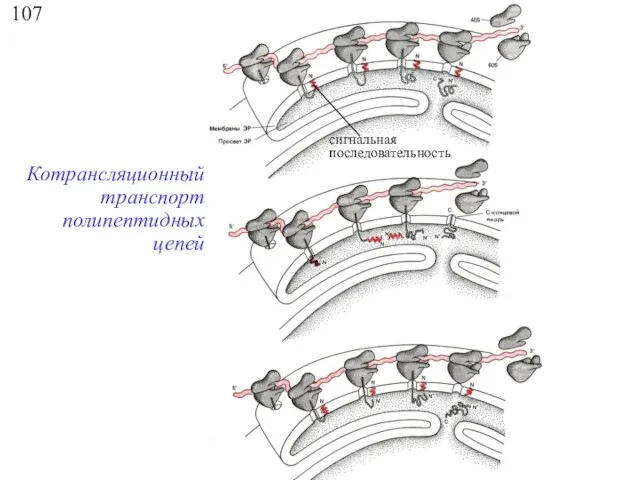
107
Котрансляционный транспорт полипептидных цепей
сигнальная
последовательность
107
Котрансляционный транспорт полипептидных цепей
сигнальная
последовательность
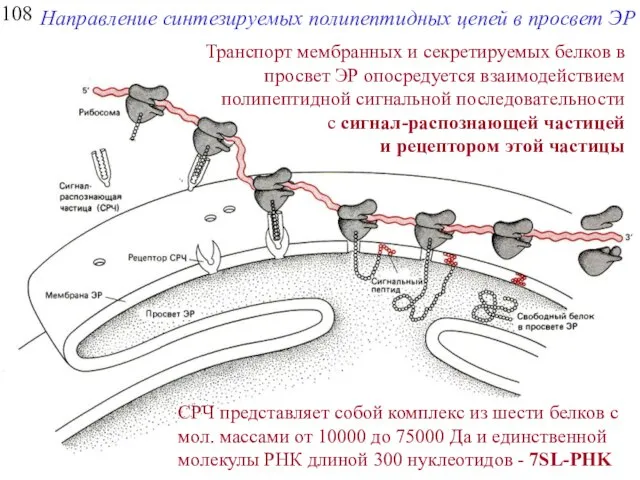
108
Направление синтезируемых полипептидных цепей в просвет ЭР
Транспорт мембранных и секретируемых белков
108
Направление синтезируемых полипептидных цепей в просвет ЭР
Транспорт мембранных и секретируемых белков
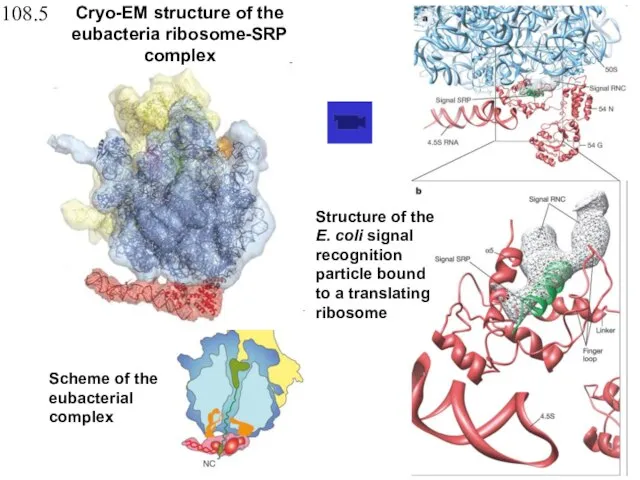
Scheme of the eubacterial complex
Structure of the E. coli signal recognition
Scheme of the eubacterial complex
Structure of the E. coli signal recognition

109
Характерные N-концевые сигнальные последовательности эукариотических и прокариотических белков
Длина сигнальной последовательности составляет
109
Характерные N-концевые сигнальные последовательности эукариотических и прокариотических белков
Длина сигнальной последовательности составляет
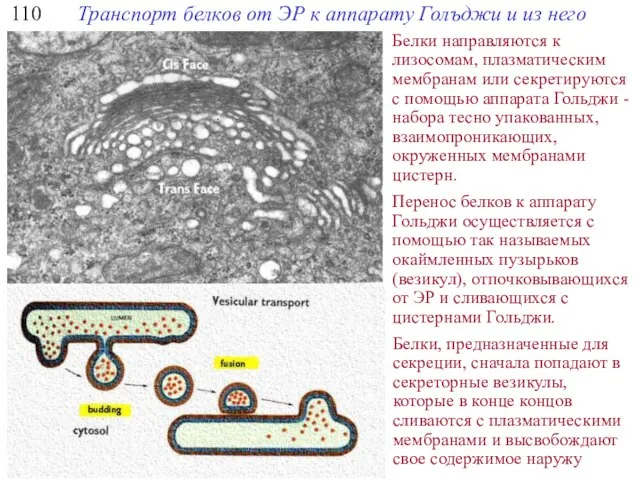
110
Транспорт белков от ЭР к аппарату Голъджи и из него
Белки направляются
110
Транспорт белков от ЭР к аппарату Голъджи и из него
Белки направляются
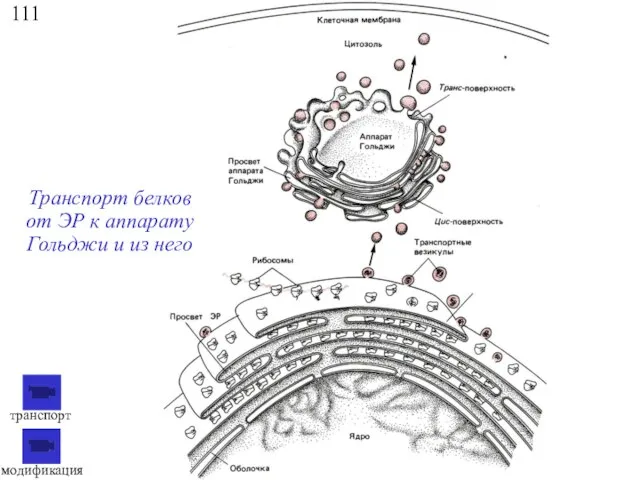
111
Транспорт белков от ЭР к аппарату Гольджи и из него
модификация
транспорт
111
Транспорт белков от ЭР к аппарату Гольджи и из него
модификация
транспорт
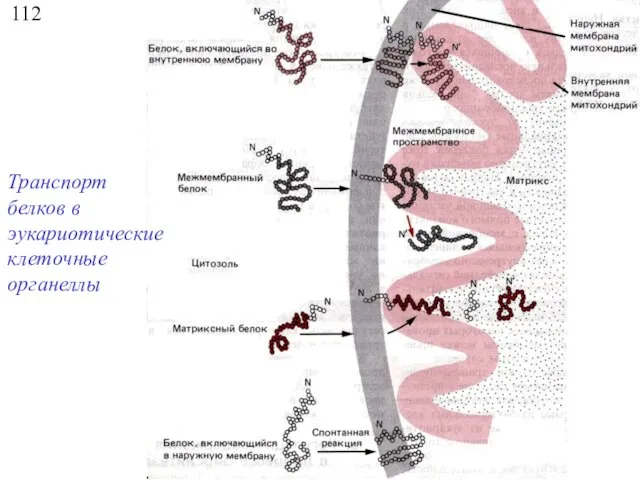
112
Транспорт белков в эукариотические клеточные органеллы
112
Транспорт белков в эукариотические клеточные органеллы
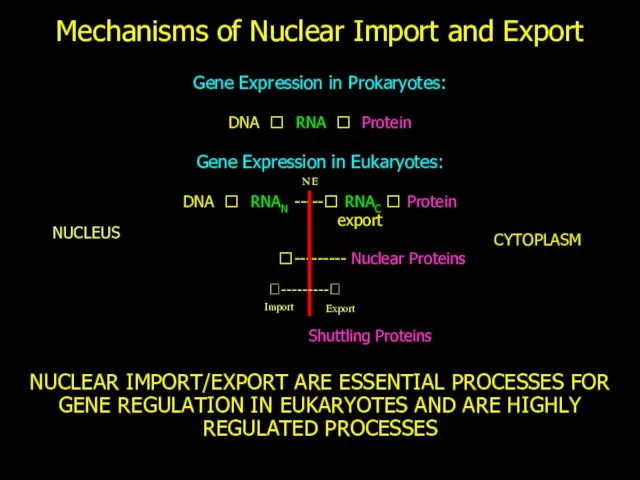
Mechanisms of Nuclear Import and Export
Gene Expression in Prokaryotes:
DNA ? RNA
Mechanisms of Nuclear Import and Export
Gene Expression in Prokaryotes:
DNA ? RNA
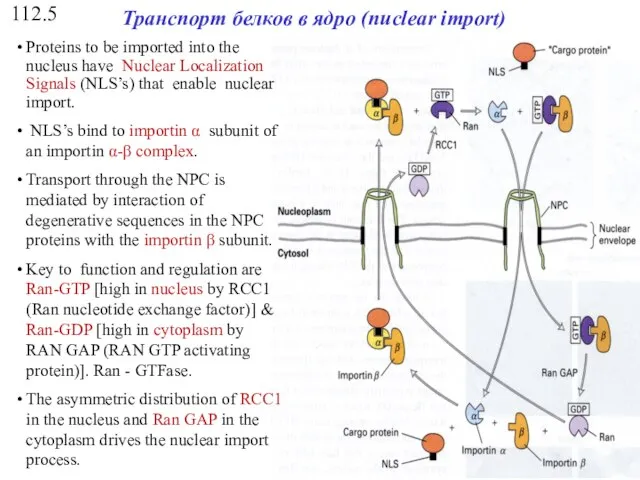
112.5
Транспорт белков в ядро (nuclear import)
Proteins to be imported into the
112.5
Транспорт белков в ядро (nuclear import)
Proteins to be imported into the
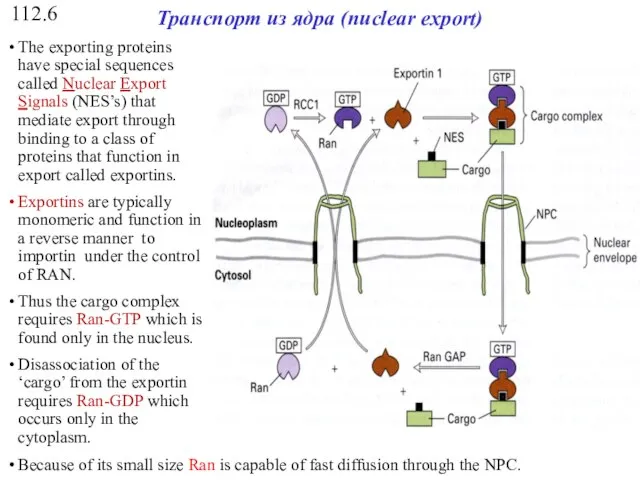
112.6
Транспорт из ядра (nuclear export)
112.6
Транспорт из ядра (nuclear export)
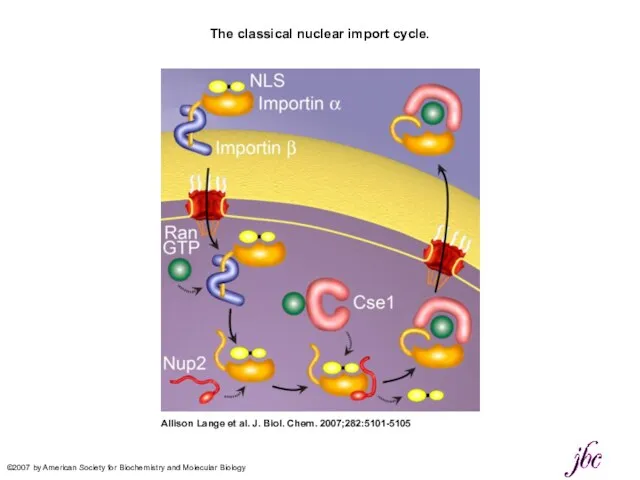
The classical nuclear import cycle.
Allison Lange et al. J. Biol.
The classical nuclear import cycle.
Allison Lange et al. J. Biol.
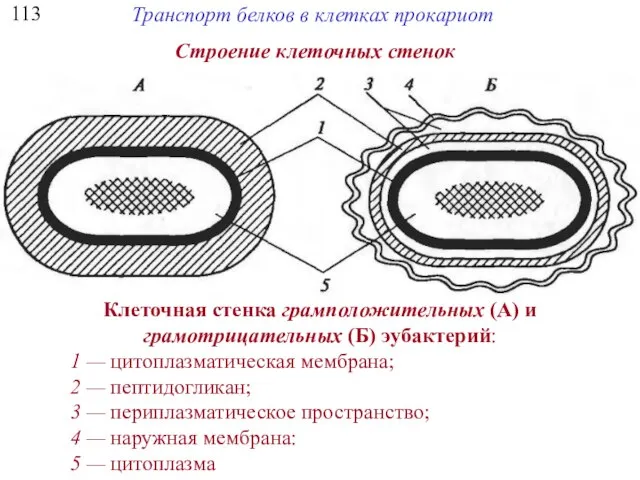
113
Клеточная стенка грамположительных (А) и
грамотрицательных (Б) эубактерий:
1 — цитоплазматическая мембрана;
113
Клеточная стенка грамположительных (А) и
грамотрицательных (Б) эубактерий:
1 — цитоплазматическая мембрана;
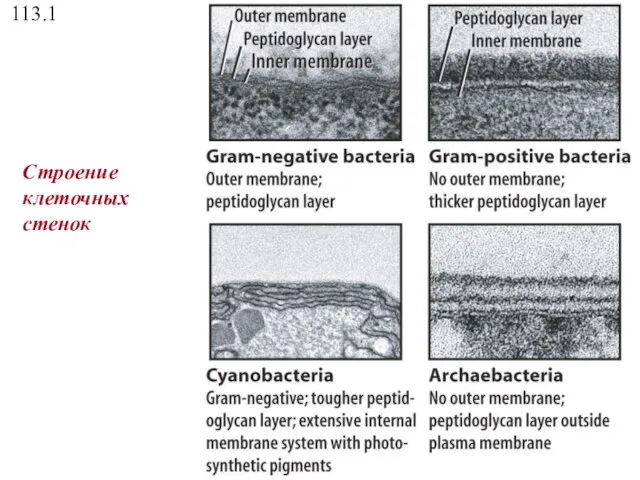
113.1
Строение клеточных стенок
113.1
Строение клеточных стенок
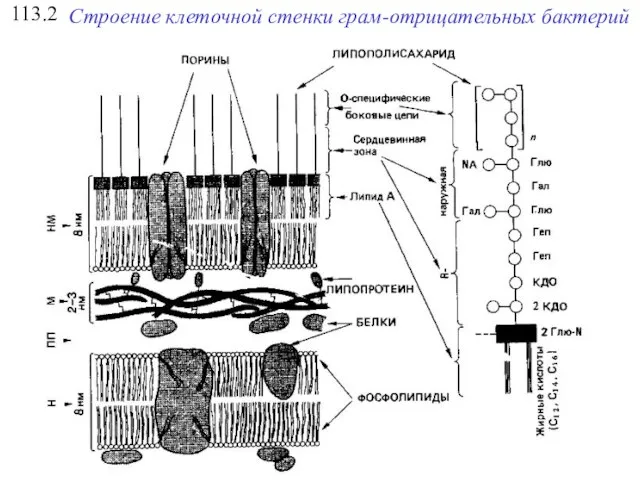
113.2
Строение клеточной стенки грам-отрицательных бактерий
113.2
Строение клеточной стенки грам-отрицательных бактерий

113.3
Строение клеточной стенки грам-положительных бактерий
113.3
Строение клеточной стенки грам-положительных бактерий

114
Характерные N-концевые сигнальные последовательности эукариотических и прокариотических белков
Длина сигнальной последовательности составляет
114
Характерные N-концевые сигнальные последовательности эукариотических и прокариотических белков
Длина сигнальной последовательности составляет

СТРУКТУРА ГЕНОМОВ
СТРУКТУРА ГЕНОМОВ

Геном - суммарная ДНК одного набора хромосом и внехромосомных генетических элементов
Геном - суммарная ДНК одного набора хромосом и внехромосомных генетических элементов
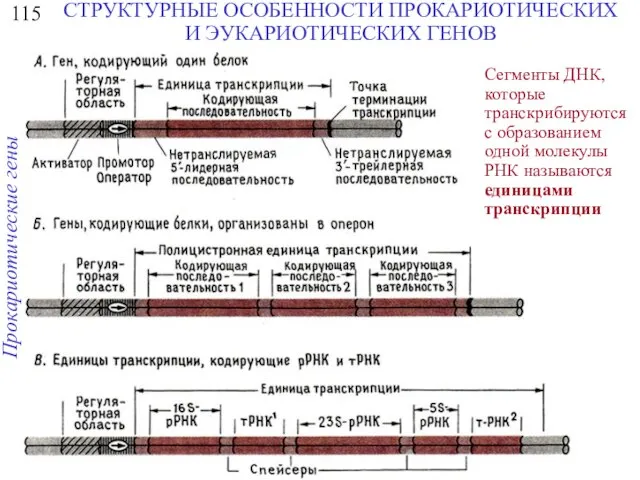
115
СТРУКТУРНЫЕ ОСОБЕННОСТИ ПРОКАРИОТИЧЕСКИХ И ЭУКАРИОТИЧЕСКИХ ГЕНОВ
Сегменты ДНК, которые транскрибируются с образованием
115
СТРУКТУРНЫЕ ОСОБЕННОСТИ ПРОКАРИОТИЧЕСКИХ И ЭУКАРИОТИЧЕСКИХ ГЕНОВ
Сегменты ДНК, которые транскрибируются с образованием
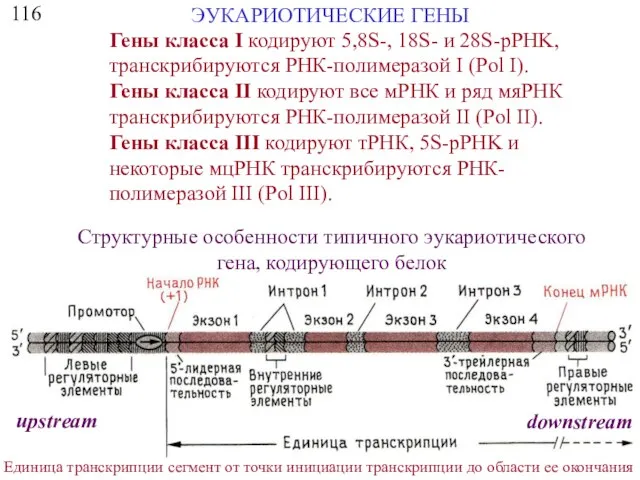
116
ЭУКАРИОТИЧЕСКИЕ ГЕНЫ
Гены класса I кодируют 5,8S-, 18S- и 28S-pPHK, транскрибируются РНК-полимеразой
116
ЭУКАРИОТИЧЕСКИЕ ГЕНЫ
Гены класса I кодируют 5,8S-, 18S- и 28S-pPHK, транскрибируются РНК-полимеразой
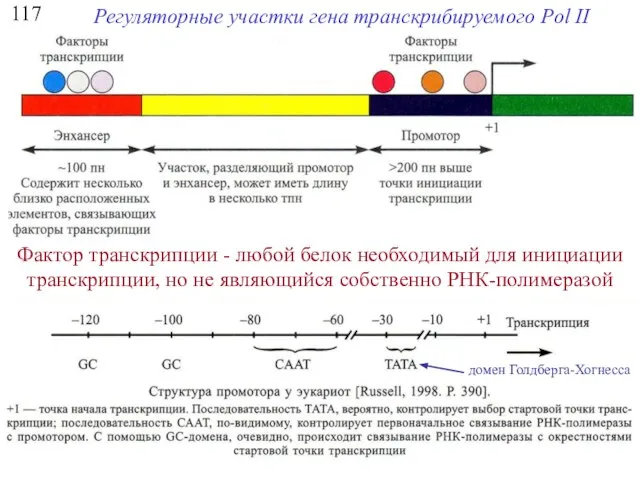
117
Регуляторные участки гена транскрибируемого Pol II
Фактор транскрипции - любой белок необходимый
117
Регуляторные участки гена транскрибируемого Pol II
Фактор транскрипции - любой белок необходимый
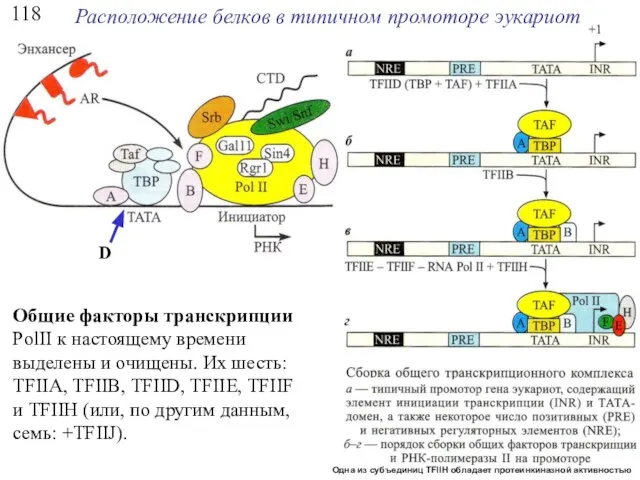
118
Расположение белков в типичном промоторе эукариот
Общие факторы транскрипции PolII к настоящему
118
Расположение белков в типичном промоторе эукариот
Общие факторы транскрипции PolII к настоящему

119
Специфические факторы транскрипции
Специфические факторы транскрипции должны обладать двумя важнейшими свойствами:
1) опознавать
119
Специфические факторы транскрипции
Специфические факторы транскрипции должны обладать двумя важнейшими свойствами:
1) опознавать
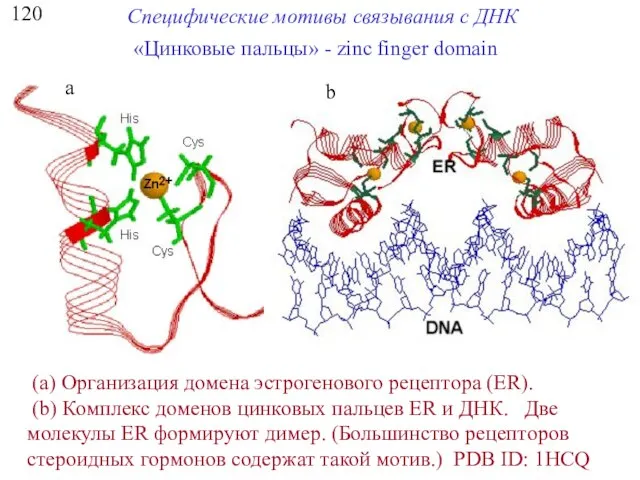
120
Специфические мотивы связывания с ДНК
«Цинковые пальцы» - zinc finger domain
120
Специфические мотивы связывания с ДНК
«Цинковые пальцы» - zinc finger domain
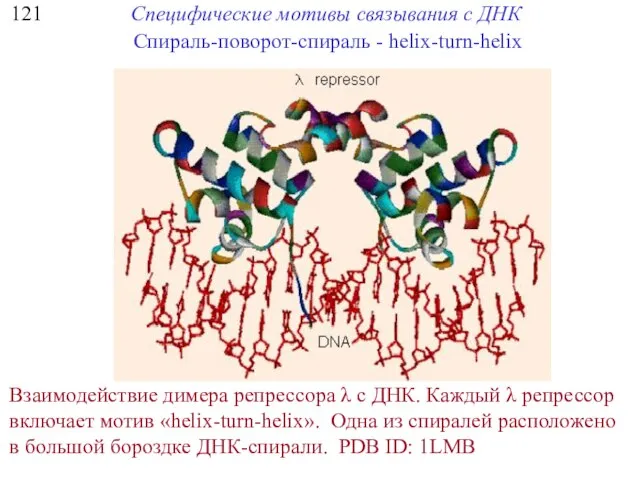
121
Специфические мотивы связывания с ДНК
Спираль-поворот-спираль - helix-turn-helix
Взаимодействие димера репрессора λ с ДНК. Каждый
121
Специфические мотивы связывания с ДНК
Спираль-поворот-спираль - helix-turn-helix
Взаимодействие димера репрессора λ с ДНК. Каждый
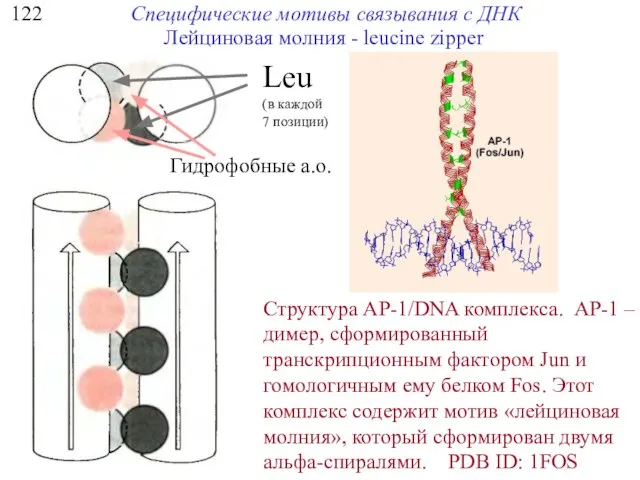
122
Специфические мотивы связывания с ДНК
Структура AP-1/DNA комплекса. AP-1 – димер, сформированный
122
Специфические мотивы связывания с ДНК
Структура AP-1/DNA комплекса. AP-1 – димер, сформированный
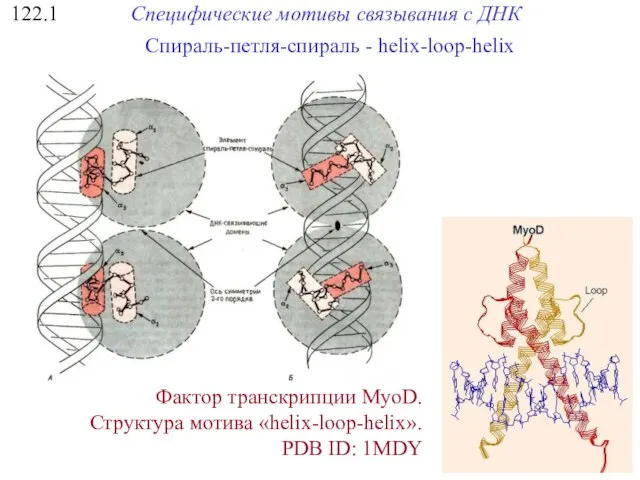
122.1
Специфические мотивы связывания с ДНК
Фактор транскрипции MyoD.
Структура мотива «helix-loop-helix».
PDB ID:
122.1
Специфические мотивы связывания с ДНК
Фактор транскрипции MyoD.
Структура мотива «helix-loop-helix».
PDB ID:
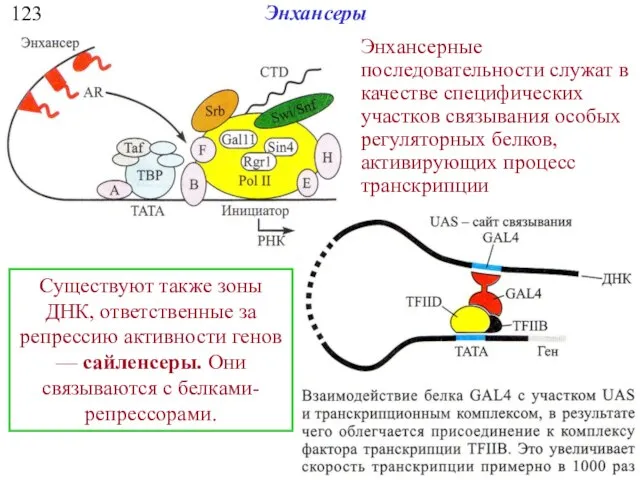
123
Энхансеры
Энхансерные последовательности служат в качестве специфических участков связывания особых регуляторных белков,
123
Энхансеры
Энхансерные последовательности служат в качестве специфических участков связывания особых регуляторных белков,
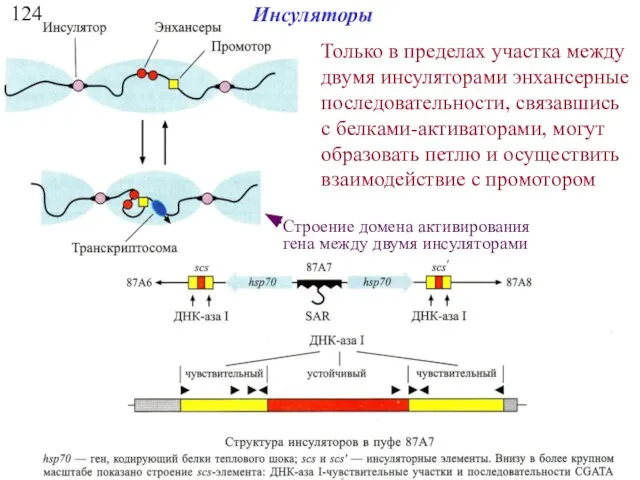
Инсуляторы
Только в пределах участка между двумя инсуляторами энхансерные последовательности, связавшись с
Инсуляторы
Только в пределах участка между двумя инсуляторами энхансерные последовательности, связавшись с

125
Терминация транскрипции
Механизмы, которые определяют 3'-конец соответствующих функциональных РНК-продуктов, уникальны для каждой
125
Терминация транскрипции
Механизмы, которые определяют 3'-конец соответствующих функциональных РНК-продуктов, уникальны для каждой
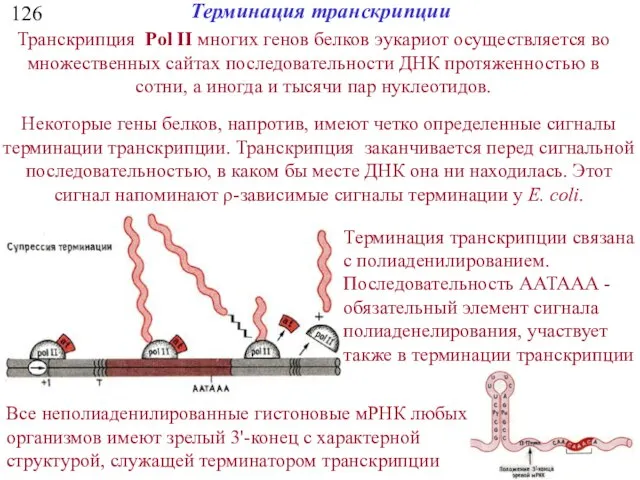
126
Транскрипция Pol II многих генов белков эукариот осуществляется во множественных сайтах
126
Транскрипция Pol II многих генов белков эукариот осуществляется во множественных сайтах
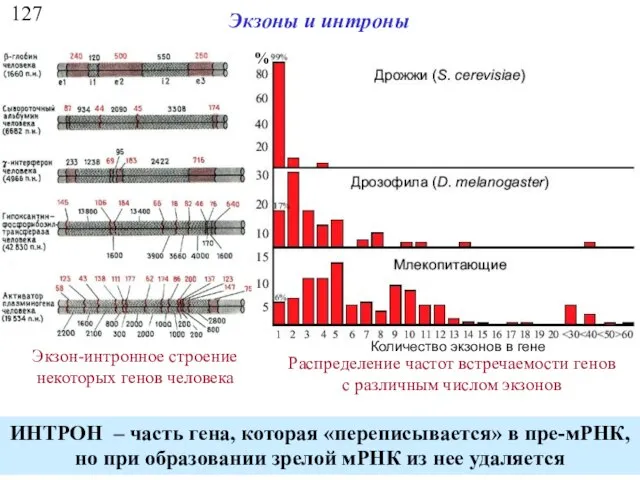
127
Экзоны и интроны
Экзон-интронное строение некоторых генов человека
Распределение частот встречаемости генов с
127
Экзоны и интроны
Экзон-интронное строение некоторых генов человека
Распределение частот встречаемости генов с
 Рациональная фармакотерапия беременных
Рациональная фармакотерапия беременных Внутриаптечный контроль лекарственных форм. Лекция № 3
Внутриаптечный контроль лекарственных форм. Лекция № 3 Неотложные состояния при инфекционных заболеваниях у детей
Неотложные состояния при инфекционных заболеваниях у детей Невербальные жесты. Жестикуляция рукой
Невербальные жесты. Жестикуляция рукой Хворий як об’єкт реабілітації
Хворий як об’єкт реабілітації Гипоспадии. Эписпадии
Гипоспадии. Эписпадии Рентгеновская дифракция – уникальный метод исследования материи
Рентгеновская дифракция – уникальный метод исследования материи Physiology of Pregnancy
Physiology of Pregnancy Психолого-педагогическая диагностика (одно из направлений деятельности специального психолога)
Психолого-педагогическая диагностика (одно из направлений деятельности специального психолога) Общая характеристика нарушений опорно-двигательного аппарата. Детский церебральный паралич
Общая характеристика нарушений опорно-двигательного аппарата. Детский церебральный паралич Индивидуально - психологические свойства личности
Индивидуально - психологические свойства личности Өмір- шексіз керуен
Өмір- шексіз керуен Медицинская приборно-компьютерная система. Разновидности МПКС. Классификация по функциональным возможностям
Медицинская приборно-компьютерная система. Разновидности МПКС. Классификация по функциональным возможностям Глоссарий (словарь терминов)
Глоссарий (словарь терминов) Туберкулез внеторакальных локализаций
Туберкулез внеторакальных локализаций Перспективы терапии Ph-негативных хронических миелопролиферативных неоплазий
Перспективы терапии Ph-негативных хронических миелопролиферативных неоплазий Изготовить лекарственную форму по прописи
Изготовить лекарственную форму по прописи Ожоговая болезнь
Ожоговая болезнь Кровотечения
Кровотечения Осуществление медицинских услуг пациентам со злокачественными новообразованиями
Осуществление медицинских услуг пациентам со злокачественными новообразованиями Сурдология. Исследование слуха
Сурдология. Исследование слуха Физиолого-гигиеническая оценка комплексного воздействия метеорологических и микроклиматических факторов
Физиолого-гигиеническая оценка комплексного воздействия метеорологических и микроклиматических факторов Мофологические элементы
Мофологические элементы Роль биологических, экологических, социальных, экономических, поведенческих факторов в формировании здоровья детей
Роль биологических, экологических, социальных, экономических, поведенческих факторов в формировании здоровья детей Дальний Восток. Финансовый результат деятельности МРФ ДВ
Дальний Восток. Финансовый результат деятельности МРФ ДВ Физиология ғылымына үлес қосқан ғалымдар
Физиология ғылымына үлес қосқан ғалымдар Современные подходы диагностике и терапии биполярного расстройства
Современные подходы диагностике и терапии биполярного расстройства Лечебные суспензии
Лечебные суспензии