- Нейровирусные инфекции

Содержание
- 2. Этиологическая структура Семейство Flaviviridae Семейство Rhabdoviridae Семейство Herpesviridae Семейство Orthomyxoviridae Семейство Paramyxoviridae Семейство Picornaviridae
- 3. Клещевой энцефалит – острая вирусная природноочаговая трансмиссивная инфекционная болезнь, характеризующаяся лихорадкой, интоксикацией и признаками поражения ЦНС.
- 4. Историческая справка Л. А. Зильбер в своей лаборатории в Институте эпидемиологии и микробиологии им. Н. Ф.
- 5. Таксономическое положение Экологическая группа арбовирусы Семейство Flaviviridae Род Flavivirus Подгруппа клещевого энцефалита
- 6. Строение вириона Вирус КЭ является сложным РНК-геномным вирусом, сферической формы, диаметром 40-60нм. Поверхностные шипы образованы Е
- 7. Белок Е и антигенные свойства вируса Белок Е – главный, биологически наиболее значимый компонент внешней поверхности
- 8. Считается, что на территории России к настоящему времени обнаружено пять антигенных вариантов (подтипов) ВКЭ: дальневосточный; западный
- 9. Генетическая вариабельность и клинические проявления «Клещевые» штаммы с большим разнообразием генофонда можно рассматривать как более гетерогенный
- 10. Эволюция вируса и перспективы Направленность эволюции клинической картины ВКЭ-инфекции во времени, за исключением дальневосточных очагов клещевого
- 11. Эволюция вируса и перспективы Основная причина наблюдаемой эволюции клещевого энцефалита заключается в природопреобразующей деятельности человека в
- 12. Геном представлен одной молекулой однонитчатой РНК положительной полярности, с короткими некодирующими областями на концах, общей длиной
- 13. Неструктурные белки Белки NS3 и NS5, по-видимому, необходимы для репликации вирусной РНК, так как в зараженных
- 14. 9. Разрыв пузырька на плазматической мембране: освобождение вириона 8. Транспорт вириона и Завершение гликозилирования 7. Сборка
- 16. Репликативный цикл В качестве специфических рецепторов ВКЭ на поверхности клеток может выступить ламининовый рецептор человека Предполагается,
- 17. Затем происходит раздевание нуклеокапсида и освобождение геномной РНК в цитоплазме. Далее следуют сложные совмещенные стадии трансляции
- 18. Клетки, инфицированные флавивирусами, кроме зрелых и дефектных вирусных частиц также секретируют медленно седиментирующие частицы с внешним
- 19. Резистентность вируса Вирус КЭ хорошо переносит низкую температуру (даже -175 С),при -150 С сохраняет жизнеспособность до
- 20. Эпидемиология клещевого энцефалита Основной резервуар и переносчик - иксодовые клещи, в особенности Ixodes persulcatus и Ixodes
- 21. Эпидемиология клещевого энцефалита (продолжение) Дополнительным природным резервуаром вируса cлужат прокормители инфицированных клещей: многочисленные грызуны, другие дикие
- 22. Эпидемиология клещевого энцефалита (продолжение) Основная масса заболеваний приходится на Уральский, Западно-Сибирский и Восточно-Сибирский регионы. В этих
- 23. Патогенез клещевого энцефалита Возбудитель проникает через кожу или слизистую ЖКТ. В месте входных ворот начинается первичная
- 24. Патогенез клещевого энцефалита (продолжение) В конце инкубационного периода в результате интенсивного размножения вирусов в эндотелии и
- 25. Патогенез клещевого энцефалита (продолжение) При остром клещевом энцефалите процессы репликативной и персистентной инфекции часто могут быть
- 26. Клиника клещевого энцефалита Острая инфекция Инкубационный период :8–23 дня (чаще 10–12 дней). Продромальный период: слабость, недомогание,
- 27. Клиника клещевого энцефалита (продолжение) Лихорадочная форма болезни характеризуется благоприятным течением с быстрым выздоровлением. Продолжительность лихорадки –
- 28. Клиника клещевого энцефалита (продолжение) Менингоэнцефалитическая очаговая форма клещевого энцефалита протекает тяжело и дает частую летальность. Больные
- 29. Клиника клещевого энцефалита (продолжение) Хроническая инфекция Представления о хроническом клещевом энцефалите обоснованы длительной персистенцией ВКЭ у
- 30. Клиника клещевого энцефалита (продолжение) Варианты исхода : Полное выздоровление с восстановлением всех функций. Сохранение или формирование
- 31. Иммунитет При остром клещевом энцефалите относительное и абсолютное содержание Т-лимфоцитов в крови снижено. Глубина вирусиндуцированной депрессии
- 32. Лабораторная диагностика клещевого энцефалита Исследуемые материалы: кровь, ликвор, испражнения, моча. Вирусологический метод. В острой стадии болезни.
- 33. Лабораторная диагностика клещевого энцефалита (продолжение) Биологический метод в острой стадии болезни основан на выделении вируса из
- 34. Спецпрофилактика и лечение Для спецпрофилактики применяют инактивированную культуральную вакцину из вируса клещевого энцефалита инактивированного формалином. Применяют
- 36. Скачать презентацию

Этиологическая структура
Семейство Flaviviridae
Семейство Rhabdoviridae
Семейство Herpesviridae
Семейство Orthomyxoviridae
Семейство Paramyxoviridae
Семейство Picornaviridae
Этиологическая структура
Семейство Flaviviridae
Семейство Rhabdoviridae
Семейство Herpesviridae
Семейство Orthomyxoviridae
Семейство Paramyxoviridae
Семейство Picornaviridae

Клещевой энцефалит – острая вирусная природноочаговая трансмиссивная инфекционная болезнь, характеризующаяся лихорадкой,
Клещевой энцефалит – острая вирусная природноочаговая трансмиссивная инфекционная болезнь, характеризующаяся лихорадкой,

Историческая справка
Л. А. Зильбер в своей лаборатории в Институте эпидемиологии
Историческая справка
Л. А. Зильбер в своей лаборатории в Институте эпидемиологии

Таксономическое положение
Экологическая группа арбовирусы
Семейство Flaviviridae
Род Flavivirus
Подгруппа клещевого энцефалита
Таксономическое положение
Экологическая группа арбовирусы
Семейство Flaviviridae
Род Flavivirus
Подгруппа клещевого энцефалита

Строение вириона
Вирус КЭ является сложным РНК-геномным вирусом, сферической формы, диаметром 40-60нм.
Строение вириона
Вирус КЭ является сложным РНК-геномным вирусом, сферической формы, диаметром 40-60нм.

Белок Е и антигенные свойства вируса
Белок Е – главный, биологически наиболее
Белок Е и антигенные свойства вируса
Белок Е – главный, биологически наиболее

Считается, что на территории России к настоящему времени обнаружено пять антигенных
Считается, что на территории России к настоящему времени обнаружено пять антигенных

Генетическая вариабельность и клинические проявления
«Клещевые» штаммы с большим разнообразием генофонда можно
Генетическая вариабельность и клинические проявления
«Клещевые» штаммы с большим разнообразием генофонда можно

Эволюция вируса и перспективы
Направленность эволюции клинической картины ВКЭ-инфекции во времени, за
Эволюция вируса и перспективы
Направленность эволюции клинической картины ВКЭ-инфекции во времени, за

Эволюция вируса и перспективы
Основная причина наблюдаемой эволюции клещевого энцефалита заключается в
Эволюция вируса и перспективы
Основная причина наблюдаемой эволюции клещевого энцефалита заключается в

Геном представлен одной молекулой однонитчатой РНК положительной полярности, с короткими некодирующими
Геном представлен одной молекулой однонитчатой РНК положительной полярности, с короткими некодирующими

Неструктурные белки
Белки NS3 и NS5, по-видимому, необходимы для репликации вирусной РНК,
Неструктурные белки
Белки NS3 и NS5, по-видимому, необходимы для репликации вирусной РНК,
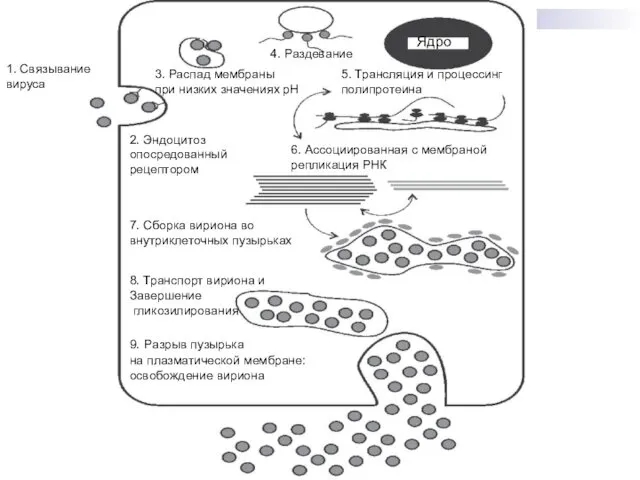
9. Разрыв пузырька
на плазматической мембране:
освобождение вириона
8. Транспорт вириона и
Завершение
гликозилирования
7. Сборка
9. Разрыв пузырька
на плазматической мембране:
освобождение вириона
8. Транспорт вириона и
Завершение
гликозилирования
7. Сборка


Репликативный цикл
В качестве специфических рецепторов ВКЭ на поверхности клеток может выступить
Репликативный цикл
В качестве специфических рецепторов ВКЭ на поверхности клеток может выступить

Затем происходит раздевание нуклеокапсида и освобождение геномной РНК в цитоплазме.
Далее следуют
Затем происходит раздевание нуклеокапсида и освобождение геномной РНК в цитоплазме.
Далее следуют

Клетки, инфицированные флавивирусами, кроме зрелых и дефектных вирусных частиц также секретируют
Клетки, инфицированные флавивирусами, кроме зрелых и дефектных вирусных частиц также секретируют

Резистентность вируса
Вирус КЭ хорошо переносит низкую температуру (даже -175 С),при -150
Резистентность вируса
Вирус КЭ хорошо переносит низкую температуру (даже -175 С),при -150
Эпидемиология клещевого энцефалита
Основной резервуар и переносчик - иксодовые клещи, в особенности
Эпидемиология клещевого энцефалита
Основной резервуар и переносчик - иксодовые клещи, в особенности
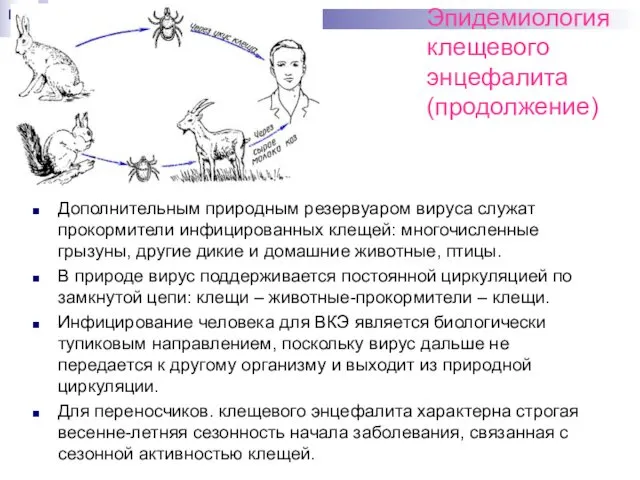
Эпидемиология клещевого
энцефалита (продолжение)
Дополнительным природным резервуаром вируса cлужат прокормители инфицированных клещей: многочисленные
Эпидемиология клещевого
энцефалита (продолжение)
Дополнительным природным резервуаром вируса cлужат прокормители инфицированных клещей: многочисленные

Эпидемиология клещевого энцефалита (продолжение)
Основная масса заболеваний приходится на Уральский, Западно-Сибирский и
Эпидемиология клещевого энцефалита (продолжение)
Основная масса заболеваний приходится на Уральский, Западно-Сибирский и

Патогенез клещевого энцефалита
Возбудитель проникает через кожу или слизистую ЖКТ. В месте
Патогенез клещевого энцефалита
Возбудитель проникает через кожу или слизистую ЖКТ. В месте

Патогенез клещевого энцефалита (продолжение)
В конце инкубационного периода в результате интенсивного размножения
Патогенез клещевого энцефалита (продолжение)
В конце инкубационного периода в результате интенсивного размножения

Патогенез клещевого энцефалита (продолжение)
При остром клещевом энцефалите процессы репликативной и персистентной
Патогенез клещевого энцефалита (продолжение)
При остром клещевом энцефалите процессы репликативной и персистентной

Клиника клещевого энцефалита
Острая инфекция
Инкубационный период :8–23 дня (чаще 10–12 дней).
Продромальный период:
Клиника клещевого энцефалита
Острая инфекция
Инкубационный период :8–23 дня (чаще 10–12 дней).
Продромальный период:

Клиника клещевого энцефалита (продолжение)
Лихорадочная форма болезни характеризуется благоприятным течением с быстрым
Клиника клещевого энцефалита (продолжение)
Лихорадочная форма болезни характеризуется благоприятным течением с быстрым

Клиника клещевого энцефалита (продолжение)
Менингоэнцефалитическая очаговая форма клещевого энцефалита протекает тяжело и
Клиника клещевого энцефалита (продолжение)
Менингоэнцефалитическая очаговая форма клещевого энцефалита протекает тяжело и

Клиника клещевого энцефалита (продолжение)
Хроническая инфекция
Представления о хроническом клещевом энцефалите обоснованы длительной
Клиника клещевого энцефалита (продолжение)
Хроническая инфекция
Представления о хроническом клещевом энцефалите обоснованы длительной

Клиника клещевого энцефалита (продолжение)
Варианты исхода :
Полное выздоровление с восстановлением
Клиника клещевого энцефалита (продолжение)
Варианты исхода :
Полное выздоровление с восстановлением

Иммунитет
При остром клещевом энцефалите относительное и абсолютное содержание Т-лимфоцитов в крови
Иммунитет
При остром клещевом энцефалите относительное и абсолютное содержание Т-лимфоцитов в крови

Лабораторная диагностика клещевого энцефалита
Исследуемые материалы: кровь, ликвор, испражнения, моча.
Вирусологический метод. В
Лабораторная диагностика клещевого энцефалита
Исследуемые материалы: кровь, ликвор, испражнения, моча.
Вирусологический метод. В

Лабораторная диагностика клещевого энцефалита (продолжение)
Биологический метод в острой стадии болезни основан
Лабораторная диагностика клещевого энцефалита (продолжение)
Биологический метод в острой стадии болезни основан

Спецпрофилактика и лечение
Для спецпрофилактики применяют инактивированную культуральную вакцину из вируса клещевого
Спецпрофилактика и лечение
Для спецпрофилактики применяют инактивированную культуральную вакцину из вируса клещевого
 Гиперандрогения (часть 2)
Гиперандрогения (часть 2) Основы лабораторной диагностики заболеваний пищеварительной системы (копрологические синдромы)
Основы лабораторной диагностики заболеваний пищеварительной системы (копрологические синдромы) Термические поражения. Ожоги и отморожения
Термические поражения. Ожоги и отморожения Конфликтный пациент с болью в спине
Конфликтный пациент с болью в спине Фармакология мочегонных, урикозурических и маточных средств
Фармакология мочегонных, урикозурических и маточных средств Роль мелатонина в репродуктивной системе женщины
Роль мелатонина в репродуктивной системе женщины Кровеносная система человека
Кровеносная система человека Федеральное казенное учреждение здравоохранения. Медико - санитарная часть
Федеральное казенное учреждение здравоохранения. Медико - санитарная часть Психические свойства личности
Психические свойства личности Сиалолитиаз. Анықтамасы. Патанатомиясы
Сиалолитиаз. Анықтамасы. Патанатомиясы Шон Стивенсон. Здоровый сон. 21 Шаг на пути к хорошему самочувствию. (2)
Шон Стивенсон. Здоровый сон. 21 Шаг на пути к хорошему самочувствию. (2) Сердечно-легочная реанимация в особых обстоятельствах
Сердечно-легочная реанимация в особых обстоятельствах Влияние курения на организм человека
Влияние курения на организм человека Общие принципы определения степени утраты профессиональной трудоспособности
Общие принципы определения степени утраты профессиональной трудоспособности Психическое развитие ребенка в младенческом возрасте
Психическое развитие ребенка в младенческом возрасте Глобальная регуляция экспрессии генов
Глобальная регуляция экспрессии генов Призвание - врач. Мазикина Людмила
Призвание - врач. Мазикина Людмила Лекарственные соединения гетероциклического ряда с одним и двумя гетероатомами
Лекарственные соединения гетероциклического ряда с одним и двумя гетероатомами Настройка параметров ИВЛ и мониторинг респираторной поддержки
Настройка параметров ИВЛ и мониторинг респираторной поддержки Голод или аппетит. Урок № 5
Голод или аппетит. Урок № 5 Профессионализм как нравственная характеристика личности
Профессионализм как нравственная характеристика личности Сахарный диабет
Сахарный диабет Кардиомиопатии
Кардиомиопатии Medical education in South Korea
Medical education in South Korea Профилактка COVID - 19
Профилактка COVID - 19 Основные гериатрические синдромы. Занятие 2
Основные гериатрические синдромы. Занятие 2 Целлюлит. Пайда болу себептері
Целлюлит. Пайда болу себептері Дуальные отношения в β-квадре в соционике
Дуальные отношения в β-квадре в соционике