- Гемоглобин и миоглобин. Протеомика. Переваривание белков. Лекция 3

Содержание
- 2. 1. Миоглобин и гемоглобин. Гемоглобин: структура, свойства, виды, возрастные изменения качественного и количественного состава в крови.
- 3. 1. Миоглобин и гемоглобин. Гемоглобин: структура, свойства, виды, возрастные изменения качественного и количественного состава в крови.
- 4. К группе гемопротеинов относятся гемоглобин и его производные, миоглобин, хлорофиллсодержащие белки и ферменты (вся цитохромная система,
- 5. Гемоглобин в качестве белкового компонента содержит глобин, а небелкового – гем. Видовые различия гемоглобина обусловлены глобином,
- 6. Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного
- 7. В молекуле гема порфин представлен в виде протопорфирина IX, содержащего четыре метильные группы (—СН3), две винильные
- 8. Модель третичной структуры молекулы миоглобина (по Дж. Кендрью). Латинскими буквами обозначены структурные домены, красным цветом –
- 9. А. Модель гемоглобина (по Перутцу). α-Цепи светлые; β-цепи темные; группы гема красные. Б. В центре молекулы
- 10. Схема строения гемоглобина Центральная полость
- 11. Строение гема, входящего в состав гемоглобина и миоглобина
- 12. Миоглобин – содержится в красных мышцах и участвует в запасании кислорода. В условиях интенсивной мышечной работы,
- 13. Гемоглобины – структурно-родственные белки, находящиеся в эритроцитах позвоночных. Они выполняют две важные биологические функции: 1) переносят
- 14. Кривые ассоциации кислорода для гемоглобина и миоглобина в зависимости от парциального давления кислорода
- 15. Наиболее распространенные гемоглобины имеют следующую тетрамерную структуру: НЬА (нормальный гемоглобин взрослого человека) – α2β2. Это основной
- 16. 2. Производные гемоглобина. Аномальные гемоглобины. Серповидно-клеточная анемия и талассемия
- 17. Мутантные (М) гемоглобины человека Мутации генов, кодирующих α- и β-цепи, могут существенным образом сказываться на их
- 18. Гемоглобин при серповидноклеточной анемии В гемоглобине S остаток Glu A2(6)β замещен на Val. Остаток А2 (Glu
- 19. Различия в строении молекул гемоглобинов 1 2 3 4 5 6 7 8 Hb A: Вал-Гис-Лей-Тре-Про-Глу-Глу-Лиз…
- 20. Этот липкий участок присутствует как в оксигенированном, так и в дезоксигенированном гемоглобине S (в гемоглобине А
- 21. Когда гемоглобин S переходит в дезоксигенированное состояние, его липкий участок связывается с комплементарным участком на другой
- 22. Ассоциация молекул дезоксигемоглобина S
- 23. Другая важная группа нарушений, связанных с аномалиями гемоглобина – талассемии. Для них характерна пониженная скорость синтеза
- 24. 3. Конформационные изменения и кооперативные взаимодействия субъединиц гемоглобина. Эффект Бора. Роль 2,3–бисфосфоглицерата
- 25. Кооперативные изменения конформации гемоглобина В дезоксигемоглобине благодаря ковалентной связи с белковой частью атом Fe2+ выступает из
- 26. Изменение положения Fe2+ и белковой части гемоглобина при присоединении О2
- 27. Кооперативные изменения конформации протомеров гемоглобина при присоединении О2
- 28. Перенос Н+ и СО2 с кровью. Эффект Бора. А – влияние концентрации СО2 и Н+ на
- 29. 2,3-Бифосфоглицерат (БФГ) – аллостерический регулятор сродства гемоглобина к О2 2,3-бисфосфоглицерат
- 30. Ткани Нb(О2)4 + БФГ ↔ Нb-БФГ + 4О2 Лёгкие
- 31. 4. Роль протеомики в оценке патологических состояний. Понятие об азотистом балансе. Переваривание и всасывание пищевых белков.
- 32. Протеомика – наука, основным предметом изучения которой являются белки, их функции и взаимодействия в живых организмах,
- 33. Основная задача протеомики – количественный анализ экспрессии белков в клетках в зависимости от их типа, состояния
- 34. Азотистый баланс означает разность между общим количеством азота, поступившим в организм человека (или другой организм), и
- 35. Если азота поступает больше, чем экскретируется, говорят, что данный индивидуум имеет положительный азотистый баланс. Примеры: период
- 36. Взрослый человек в норме находится в состоянии азотистого равновесия: потребление азота уравновешивается его выделением в составе
- 37. В состоянии отрицательного азотистого баланса количество выделяемого азота превышает количество азота, потребляемого организмом. Важным примером служат
- 38. Переваривание пищевых белков Переваривание белков, входящих в состав пищи, происходит в различных отделах ЖКТ с участием
- 39. Пепсин Основная пищеварительная функция желудка заключается в том, что в нем начинается переваривание белка. Пепсин продуцируется
- 40. Пепсин преобразует денатурированный белок в протеозы и затем в пептоны —большие полипептидные производные. Он представляет собой
- 41. Гастриксин – протеолитический фермент (катализирующий расщепление белка пищи) желудочного сока, проявляющий максимальную активность при pH, равном
- 42. Гастриксин, в отличие от пепсина, протеолетически активен в менее кислой среде. Гастриксин является вторым по объёму
- 43. Реннин (химозин, сычужный фермент). Этот фермент вызывает створаживание молока. Он чрезвычайно важен для процессов пищеварения у
- 44. Трипсин, химотрипсин и эластаза Протеолитическое действие панкреатического секрета обусловливается тремя эндопептидазами – трипсином, химотрипсином и эластазой,
- 45. Все три фермента секретируются в виде зимогенов. Активация трипсиногена осуществляется другим протеолитическим ферментом, энтерокиназой, секретируемой слизистой
- 46. Карбоксипептидаза Дальнейшее расщепление полипептидов, образовавшихся под действием эндопептидаз, осуществляет экзопептидаза – карбоксипептидаза, которая атакует С-концевую пептидную
- 47. Переваривание секретом кишечника Кишечный сок, секретируемый железами Бруннера и Либеркюна, также содержит пищеварительные ферменты, в число
- 48. Переваривание секретом кишечника 3) фосфатаза, удаляющая фосфат из некоторых органических фосфатов (гексозофосфаты и глицерофосфат) и из
- 49. Всасывание аминокислот и белка В нормальных условиях пищевые белки почти полностью расщепляются на составляющие их аминокислоты,
- 50. Изменения белкового состава организма Изменение белкового состава клеток в процессе их дифференцировки. Наследственные протеинопатии. Приобретённые протеинопатии.
- 51. 5. Бактериальное расщепление невсосавшихся в кишечнике аминокислот
- 52. Процессы гниения и брожения в кишечнике Большая часть потребленной пищи всасывается в тонком кишечнике. Остальная часть
- 53. Судьба аминокислот в кишечнике Большинство аминокислот подвергается декарбоксилированию в результате действия кишечных бактерий с образованием токсических
- 54. [2H] CH3SH → CH4 + H2S Метилмеркаптан Метан Сероводород Аминокислота триптофан в результате нескольких реакций превращается
- 55. Превращение аминокислот в толстом кишечнике
- 57. Скачать презентацию

1. Миоглобин и гемоглобин. Гемоглобин: структура, свойства, виды, возрастные изменения качественного
1. Миоглобин и гемоглобин. Гемоглобин: структура, свойства, виды, возрастные изменения качественного

1. Миоглобин и гемоглобин. Гемоглобин: структура, свойства, виды, возрастные изменения качественного
1. Миоглобин и гемоглобин. Гемоглобин: структура, свойства, виды, возрастные изменения качественного

К группе гемопротеинов относятся гемоглобин и его производные, миоглобин, хлорофиллсодержащие белки
К группе гемопротеинов относятся гемоглобин и его производные, миоглобин, хлорофиллсодержащие белки

Гемоглобин в качестве белкового компонента содержит глобин, а небелкового – гем.
Гемоглобин в качестве белкового компонента содержит глобин, а небелкового – гем.

Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся
Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся

В молекуле гема порфин представлен в виде протопорфирина IX, содержащего четыре
В молекуле гема порфин представлен в виде протопорфирина IX, содержащего четыре
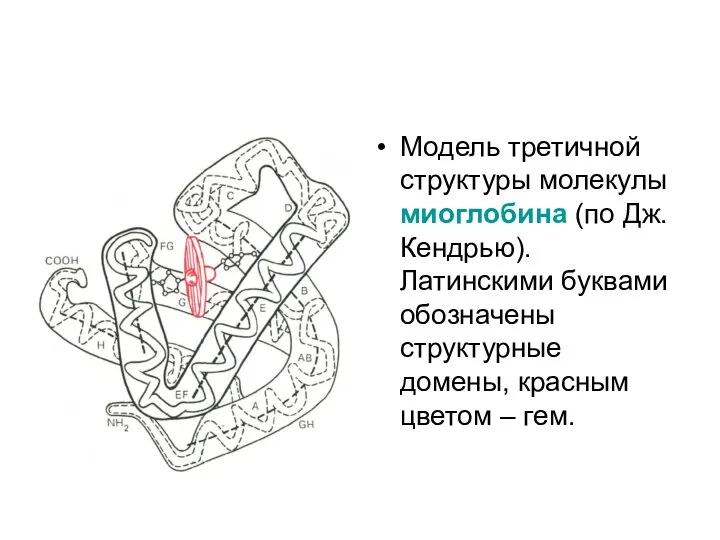
Модель третичной структуры молекулы миоглобина (по Дж. Кендрью). Латинскими буквами обозначены
Модель третичной структуры молекулы миоглобина (по Дж. Кендрью). Латинскими буквами обозначены
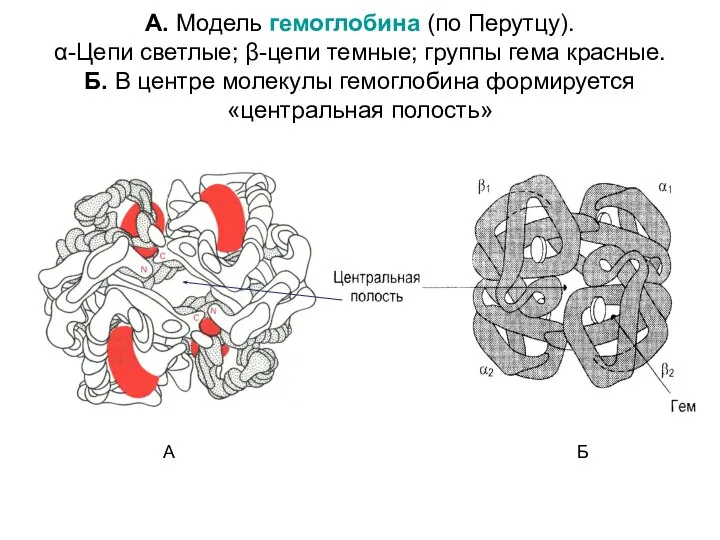
А. Модель гемоглобина (по Перутцу).
α-Цепи светлые; β-цепи темные; группы гема
А. Модель гемоглобина (по Перутцу). α-Цепи светлые; β-цепи темные; группы гема
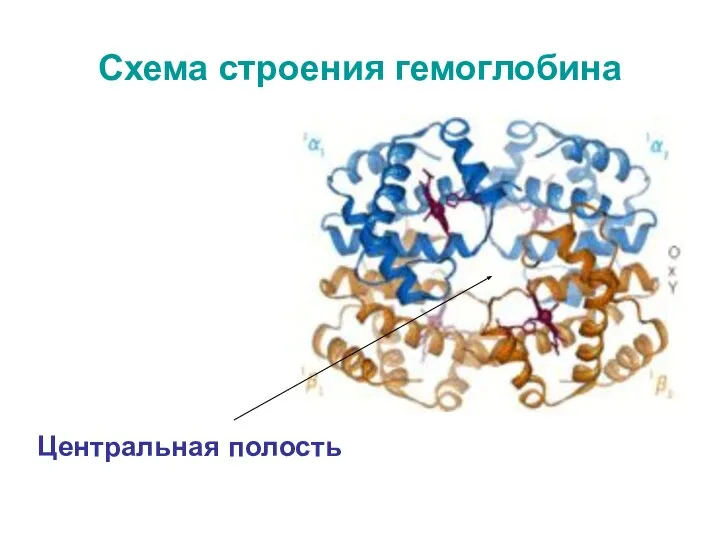
Схема строения гемоглобина
Центральная полость
Схема строения гемоглобина
Центральная полость
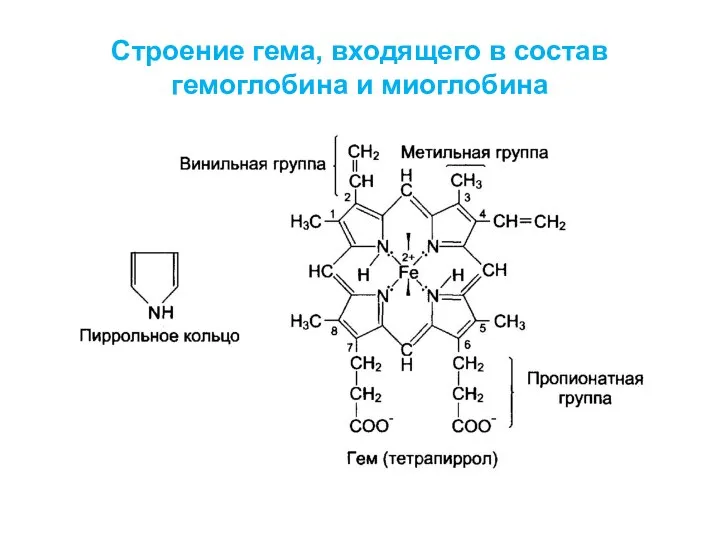
Строение гема, входящего в состав гемоглобина и миоглобина
Строение гема, входящего в состав гемоглобина и миоглобина

Миоглобин – содержится в красных мышцах и участвует в запасании кислорода.
Миоглобин – содержится в красных мышцах и участвует в запасании кислорода.

Гемоглобины – структурно-родственные белки, находящиеся в эритроцитах позвоночных. Они выполняют две
Гемоглобины – структурно-родственные белки, находящиеся в эритроцитах позвоночных. Они выполняют две
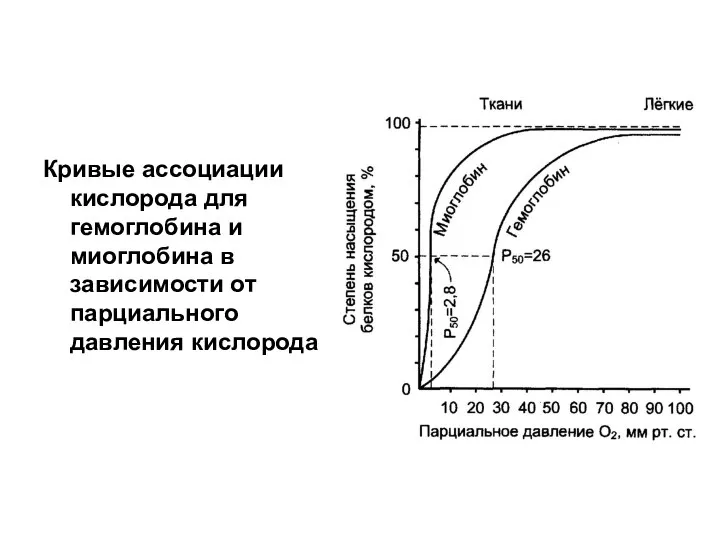
Кривые ассоциации кислорода для гемоглобина и миоглобина в зависимости от парциального

Наиболее распространенные гемоглобины имеют следующую тетрамерную структуру:
НЬА (нормальный гемоглобин взрослого
Наиболее распространенные гемоглобины имеют следующую тетрамерную структуру:
НЬА (нормальный гемоглобин взрослого

2. Производные гемоглобина. Аномальные гемоглобины. Серповидно-клеточная анемия и талассемия
2. Производные гемоглобина. Аномальные гемоглобины. Серповидно-клеточная анемия и талассемия

Мутантные (М) гемоглобины человека
Мутации генов, кодирующих α- и β-цепи, могут существенным
Мутантные (М) гемоглобины человека
Мутации генов, кодирующих α- и β-цепи, могут существенным

Гемоглобин при серповидноклеточной анемии
В гемоглобине S остаток Glu A2(6)β замещен на
Гемоглобин при серповидноклеточной анемии
В гемоглобине S остаток Glu A2(6)β замещен на

Различия в строении молекул гемоглобинов
1 2 3 4 5 6
Различия в строении молекул гемоглобинов
1 2 3 4 5 6

Этот липкий участок присутствует как в оксигенированном, так и в дезоксигенированном
Этот липкий участок присутствует как в оксигенированном, так и в дезоксигенированном

Когда гемоглобин S переходит в дезоксигенированное состояние, его липкий участок связывается
Когда гемоглобин S переходит в дезоксигенированное состояние, его липкий участок связывается

Ассоциация молекул дезоксигемоглобина S
Ассоциация молекул дезоксигемоглобина S

Другая важная группа нарушений, связанных с аномалиями гемоглобина – талассемии.
Для
Другая важная группа нарушений, связанных с аномалиями гемоглобина – талассемии.
Для

3. Конформационные изменения и кооперативные взаимодействия субъединиц гемоглобина. Эффект Бора. Роль
3. Конформационные изменения и кооперативные взаимодействия субъединиц гемоглобина. Эффект Бора. Роль

Кооперативные изменения конформации гемоглобина
В дезоксигемоглобине благодаря ковалентной связи с белковой частью
Кооперативные изменения конформации гемоглобина
В дезоксигемоглобине благодаря ковалентной связи с белковой частью
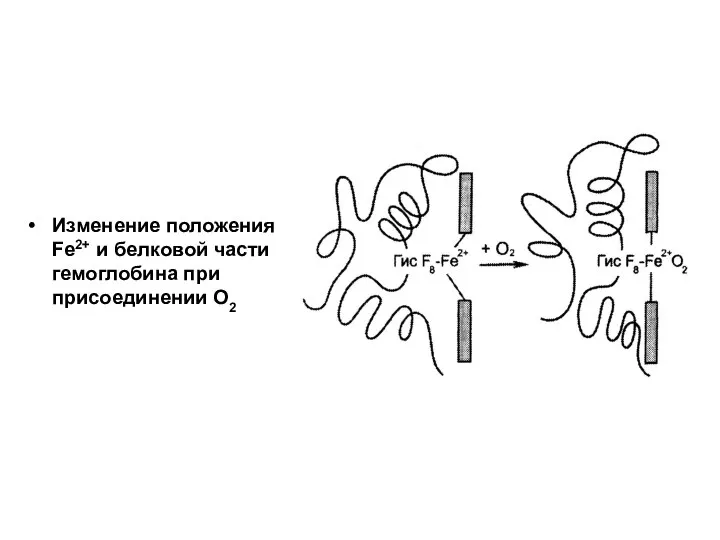
Изменение положения Fe2+ и белковой части гемоглобина при присоединении О2
Изменение положения Fe2+ и белковой части гемоглобина при присоединении О2
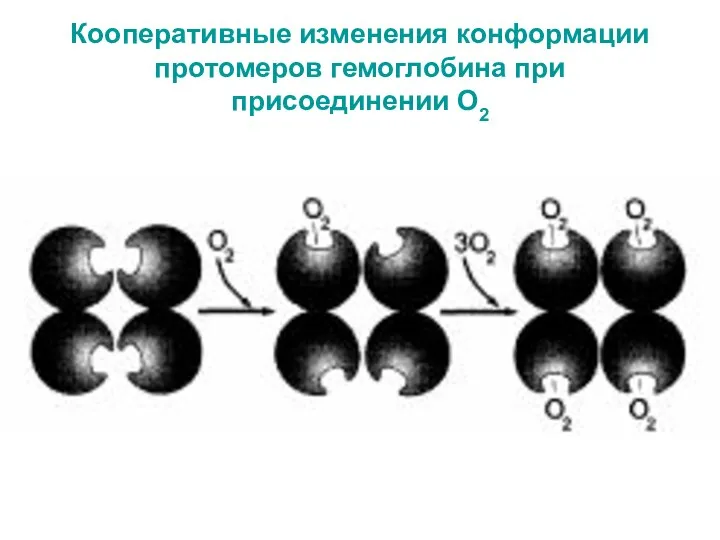
Кооперативные изменения конформации протомеров гемоглобина при присоединении О2
Кооперативные изменения конформации протомеров гемоглобина при присоединении О2
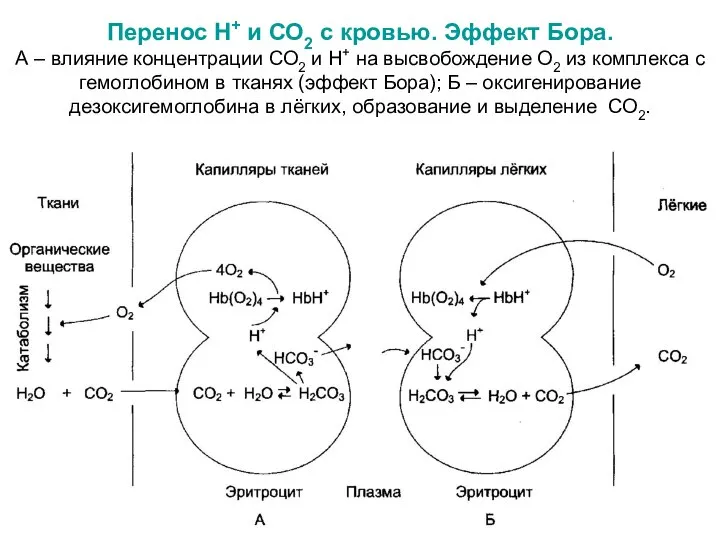
Перенос Н+ и СО2 с кровью. Эффект Бора.
А – влияние концентрации
Перенос Н+ и СО2 с кровью. Эффект Бора. А – влияние концентрации
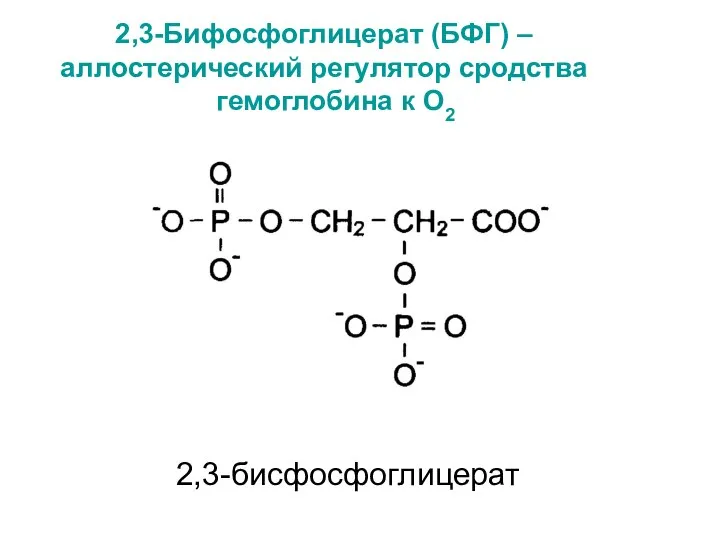
2,3-Бифосфоглицерат (БФГ) – аллостерический регулятор сродства гемоглобина к О2
2,3-бисфосфоглицерат
2,3-Бифосфоглицерат (БФГ) – аллостерический регулятор сродства гемоглобина к О2
2,3-бисфосфоглицерат
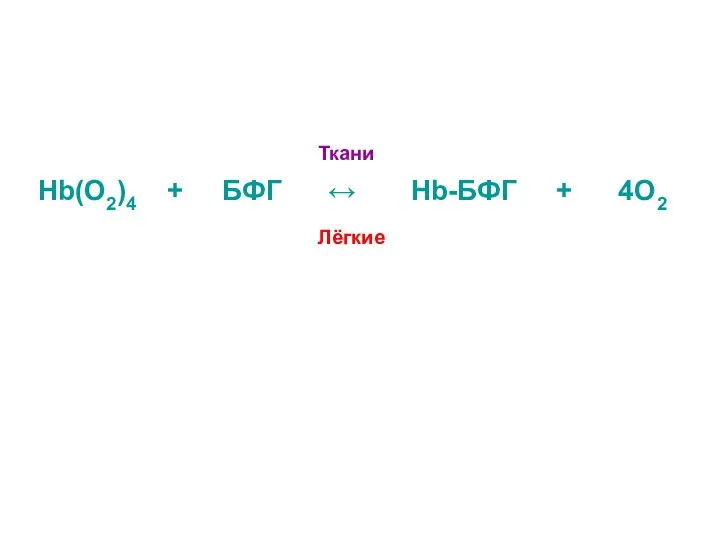
Ткани
Нb(О2)4 + БФГ ↔ Нb-БФГ + 4О2
Лёгкие
Ткани
Нb(О2)4 + БФГ ↔ Нb-БФГ + 4О2
Лёгкие

4. Роль протеомики в оценке патологических состояний. Понятие об азотистом балансе.
4. Роль протеомики в оценке патологических состояний. Понятие об азотистом балансе.

Протеомика – наука, основным предметом изучения которой являются белки, их функции
Протеомика – наука, основным предметом изучения которой являются белки, их функции

Основная задача протеомики – количественный анализ экспрессии белков в клетках в
Основная задача протеомики – количественный анализ экспрессии белков в клетках в

Азотистый баланс означает разность между общим количеством азота, поступившим в организм
Азотистый баланс означает разность между общим количеством азота, поступившим в организм

Если азота поступает больше, чем экскретируется, говорят, что данный индивидуум имеет
Если азота поступает больше, чем экскретируется, говорят, что данный индивидуум имеет

Взрослый человек в норме находится в состоянии азотистого равновесия: потребление азота
Взрослый человек в норме находится в состоянии азотистого равновесия: потребление азота

В состоянии отрицательного азотистого баланса количество выделяемого азота превышает количество азота,
В состоянии отрицательного азотистого баланса количество выделяемого азота превышает количество азота,

Переваривание пищевых белков
Переваривание белков, входящих в состав пищи, происходит в различных
Переваривание пищевых белков
Переваривание белков, входящих в состав пищи, происходит в различных

Пепсин
Основная пищеварительная функция желудка заключается в том, что в нем начинается
Пепсин
Основная пищеварительная функция желудка заключается в том, что в нем начинается

Пепсин преобразует денатурированный белок в протеозы и затем в пептоны —большие
Пепсин преобразует денатурированный белок в протеозы и затем в пептоны —большие

Гастриксин – протеолитический фермент (катализирующий расщепление белка пищи) желудочного сока, проявляющий
Гастриксин – протеолитический фермент (катализирующий расщепление белка пищи) желудочного сока, проявляющий

Гастриксин, в отличие от пепсина, протеолетически активен в менее кислой среде.
Гастриксин, в отличие от пепсина, протеолетически активен в менее кислой среде.

Реннин (химозин, сычужный фермент). Этот фермент вызывает створаживание молока. Он чрезвычайно
Реннин (химозин, сычужный фермент). Этот фермент вызывает створаживание молока. Он чрезвычайно

Трипсин, химотрипсин и эластаза
Протеолитическое действие панкреатического секрета обусловливается тремя эндопептидазами –
Трипсин, химотрипсин и эластаза
Протеолитическое действие панкреатического секрета обусловливается тремя эндопептидазами –

Все три фермента секретируются в виде зимогенов. Активация трипсиногена осуществляется другим
Все три фермента секретируются в виде зимогенов. Активация трипсиногена осуществляется другим

Карбоксипептидаза
Дальнейшее расщепление полипептидов, образовавшихся под действием эндопептидаз, осуществляет экзопептидаза – карбоксипептидаза,
Карбоксипептидаза
Дальнейшее расщепление полипептидов, образовавшихся под действием эндопептидаз, осуществляет экзопептидаза – карбоксипептидаза,

Переваривание секретом кишечника
Кишечный сок, секретируемый железами Бруннера и Либеркюна, также содержит
Переваривание секретом кишечника
Кишечный сок, секретируемый железами Бруннера и Либеркюна, также содержит

Переваривание секретом кишечника
3) фосфатаза, удаляющая фосфат из некоторых органических фосфатов (гексозофосфаты
Переваривание секретом кишечника
3) фосфатаза, удаляющая фосфат из некоторых органических фосфатов (гексозофосфаты

Всасывание аминокислот и белка
В нормальных условиях пищевые белки почти полностью расщепляются
Всасывание аминокислот и белка
В нормальных условиях пищевые белки почти полностью расщепляются

Изменения белкового состава организма
Изменение белкового состава клеток в процессе их дифференцировки.
Наследственные
Изменения белкового состава организма
Изменение белкового состава клеток в процессе их дифференцировки.
Наследственные

5. Бактериальное расщепление невсосавшихся в кишечнике аминокислот
5. Бактериальное расщепление невсосавшихся в кишечнике аминокислот

Процессы гниения и брожения в кишечнике
Большая часть потребленной пищи всасывается в
Процессы гниения и брожения в кишечнике
Большая часть потребленной пищи всасывается в
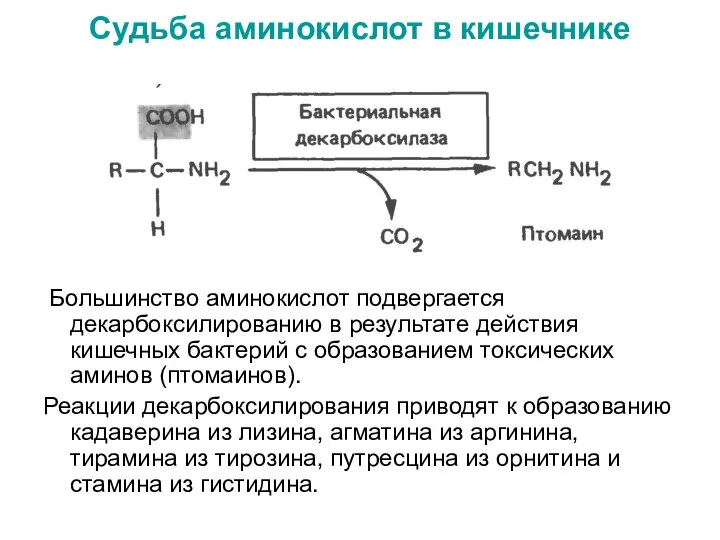
Судьба аминокислот в кишечнике
Большинство аминокислот подвергается декарбоксилированию в результате действия
Судьба аминокислот в кишечнике
Большинство аминокислот подвергается декарбоксилированию в результате действия
![[2H] CH3SH → CH4 + H2S Метилмеркаптан Метан Сероводород Аминокислота триптофан](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/749528/slide-53.jpg)
[2H]
CH3SH → CH4 + H2S Метилмеркаптан Метан Сероводород
Аминокислота триптофан
[2H]
CH3SH → CH4 + H2S Метилмеркаптан Метан Сероводород
Аминокислота триптофан
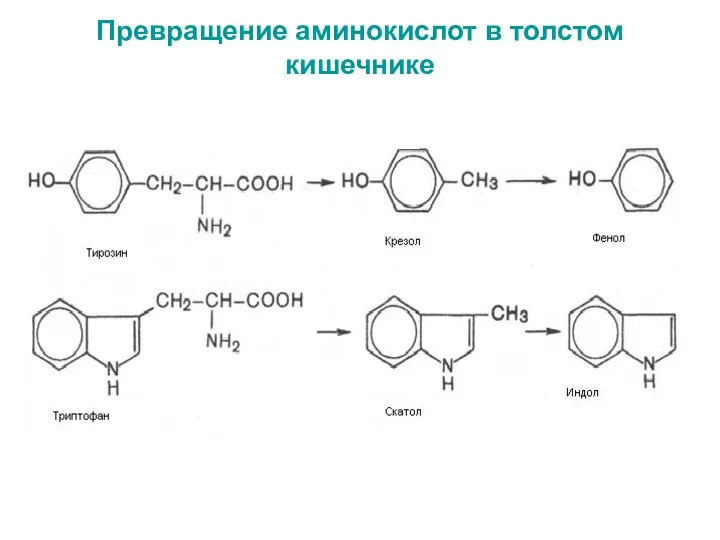
Превращение аминокислот в толстом кишечнике
Превращение аминокислот в толстом кишечнике
 Белый медведь Кулькова Ольга
Белый медведь Кулькова Ольга Основные методы селекции животных
Основные методы селекции животных Понятие о природном сообществе, биогеоценозе и экосистеме. Приспособленность растений к совместной жизни в природном сообществе
Понятие о природном сообществе, биогеоценозе и экосистеме. Приспособленность растений к совместной жизни в природном сообществе ДНК. РНК. АТФ
ДНК. РНК. АТФ Генетика человека
Генетика человека Сенсорные возможности животных
Сенсорные возможности животных Муниципальное бюджетное образовательное учреждение Средняя общеобразовательная школа № 121 .. «ПОЧЕМУ ЖИВЫЕ ОРГАНИЗМЫ НЕ РАС
Муниципальное бюджетное образовательное учреждение Средняя общеобразовательная школа № 121 .. «ПОЧЕМУ ЖИВЫЕ ОРГАНИЗМЫ НЕ РАС Императорские пингвины
Императорские пингвины  Обитатели садов и огородов
Обитатели садов и огородов Плесневые грибы и дрожжи
Плесневые грибы и дрожжи Способы питания животных.
Способы питания животных. Особенности морфологии и строения других групп микроорганизмов
Особенности морфологии и строения других групп микроорганизмов Африканская саванна
Африканская саванна Высшая нервная деятельность
Высшая нервная деятельность Выявления факторов, ограничивающих рост и развитие растений
Выявления факторов, ограничивающих рост и развитие растений Экологические группы гидробионтов.
Экологические группы гидробионтов. Урок-игра по теме: «Путешествие по эукариотической клетке» Разработала учитель биологии МОУ СОШ №18 Города Костромы Рыбкина На
Урок-игра по теме: «Путешествие по эукариотической клетке» Разработала учитель биологии МОУ СОШ №18 Города Костромы Рыбкина На Оленячі роги та їхне застосування
Оленячі роги та їхне застосування Лабораторная работа «Одноклеточные и многоклеточные организмы под микроскопом»
Лабораторная работа «Одноклеточные и многоклеточные организмы под микроскопом» Исследовательский проект Кактус – колючий друг
Исследовательский проект Кактус – колючий друг Социально-биологические основы физической культуры
Социально-биологические основы физической культуры растения поля
растения поля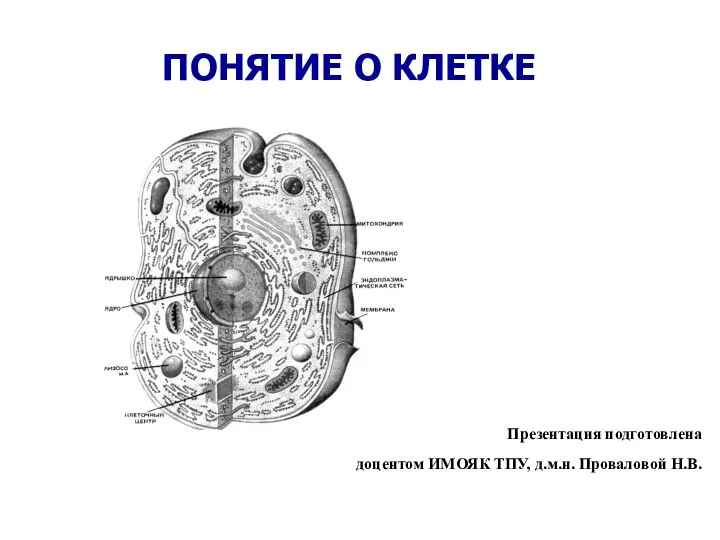 ПОНЯТИЕ О КЛЕТКЕ
ПОНЯТИЕ О КЛЕТКЕ Критерий согласия. Практический пример применения критерия согласия. Закон Менделя
Критерий согласия. Практический пример применения критерия согласия. Закон Менделя Уссурийские тигры. Научно-практическая конференция Взгляд в будущее-2017. Проект их жизнь в наших руках
Уссурийские тигры. Научно-практическая конференция Взгляд в будущее-2017. Проект их жизнь в наших руках Презентация на тему Строение и функции пищеварительной системы
Презентация на тему Строение и функции пищеварительной системы Флора и Фауна Кубани.
Флора и Фауна Кубани. Фізіологія збудливих тканин
Фізіологія збудливих тканин