- Биохимия эритроцитов, гемоглобина и лейкоцитов. Кровь. (Лекция 17)

Содержание
- 2. План лекции Эритроцит: особенности структуры, химического состава, метаболизма белков, жиров, углеводов. Гемоглобин и миоглобин: строение, функции,
- 3. Эритроциты форменные элементы крови образуются в костном мозге, циркулируют в крови около 100-120 дней, разрушаются макрофагами
- 5. ФУНКЦИИ ЭРИТРОЦИТОВ Газообмен: транспорт О2 и СО2 Регуляция КОС Адсорбция и транспорт на клеточной мембране различных
- 6. Строение эритроцита 60 % вода 40% сухой остаток 5 % - органические и неорганические вещества 95%
- 7. II. Плазмолемма Белки (15 шт): Спектрин Актин Белок 3 полосы Гликофорин Белок полосы 4.1 Анкирин Na+/K+-АТФ-аза
- 8. Часть белков плазмолеммы образуют цитоскелет фибриллярный белок спектрин образует сеть глобулярные белки, пронизывающие мембрану: гликофорин ,
- 9. олигосахарид Cl- HCO3-
- 11. Спектрин основной белок цитоскелета эритроцитов ( 25% массы всех белков), имеет вид фибриллы 100нм, состоящей из
- 12. Гликофорин трансмембранный гликопротеин (30 кДа), пронизывает плазмолемму в виде спирали. С наружной поверхности эритроцита к нему
- 13. Углеводы олигосахариды гликолипидов и гликопротеидов, на внешней поверхности плазмолеммы образуют гликокаликс. Олигосахариды гликофорина являются агглютиногенами (А
- 14. ОСОБЕННОСТЬ ОБМЕНА ВЕЩЕСТВ И ЭНЕРГИИ В ЭРИТРОЦИТЕ Эритроцит высокоспециализированная клетка, хорошо приспособленная для транспорта газов. Для
- 15. Особенности обмена нуклеотидов рибозо-5ф + аденин → АМФ АМФ + АТФ → АДФ + АДФ АДФ
- 16. Особенности обмена углеводов Анаэробный гликолиз ПФШ 2,3 дифосфоглице- ратный шунт Глюкоза Глюкоза Глюкозо-6ф ГЛЮТ-1 инсулиннезависим Гексокиназа
- 17. (метод оценки ПРЭ)
- 18. Обмен метгемоглобина В течение суток до 3% гемоглобина может спонтанно окисляться в метгемоглобин: Ηb (Fe2+) →
- 19. Восстановление метгемоглобина также может осуществляться неферментативным путём за счёт витамина В12, аскорбиновой кислоты или глутатиона Генетический
- 20. ОБРАЗОВАНИЕ ТЕЛЕЦ ХА ПРИ СЕРПОВИДНОКЛЕТОЧ НОЙ АНЕМИИ
- 21. Na+/K+-АТФ-аза - мембранный фермент поддерживает градиент концентраци Na+ и К+ Патология (сфероцитоз): ↓Na+/K+-АТФ-азы → ↑ Na+
- 23. Особенности липидного обмена В зрелом эритроците липиды не синтезируются, эритроцит может обмениваться липидами с липопротеинами крови.
- 24. В зрелом эритроците белки не синтезируются, т.к. у него нет рибосом, но в цитоплазме синтезируется глутатион
- 25. Структура трипептида глутатиона (L-глутамил-L-цистеинилглицин, GSH)
- 26. Биосинтез глутатиона Глутатион синтезируется из аминокислот L-цистеина, L-глутаминовой кислоты и глицина. Синтез происходит в две АТР-зависимые
- 27. Особенность метаболизма при старении эритроцитов ↓ образование АТФ Нарушаются энергозависимые процессы восстановления формы эритроцитов → сфероцитоз,
- 28. СТРОЕНИЕ ГЕМА Гем - это порфирин, в центре которого находиться Fe2+. Fe2+ включается в молекулу порфирина
- 29. Наибольшее количество гема содержат эритроциты (Hb), мышечные клетки (миоглобин) и клетки печени (цитохром Р450). Гемы разных
- 30. СИНТЕЗ ГЕМА Гем синтезируется во всех тканях, но с наибольшей скоростью в костном мозге и печени.
- 31. В цитоплазме Гидроксиметилбилан Уропорфириноген III Копропорфириноген III Протопорфириноген IX Протопорфирин IX Гем Порфобилиногендезаминаза Уропорфириноген III косинтаза
- 32. Порфирии - гетерогенная группа заболеваний, вызванная нарушениями синтеза гема вследствие дефицита одного или нескольких ферментов. НАРУШЕНИЯ
- 34. Негативные последствия порфирий связаны с дефицитом гема и накоплением в организме промежуточных продуктов синтеза гема –
- 35. В коже на солнце порфириногены превращаются в порфирины. О2 при взаимодействии с порфиринами переходит в синглетное
- 36. Порфириногены бесцветны и не флуоресцируют, а порфирины имеют интенсивную красную флуоресценцию в ультрафиолетовых лучах. Избыток порфиринов
- 37. Гемоглобин Тетрамерный хромопротеин 4 гема и 4 глобина (α, β, γ, δ и т.д.) Масса –
- 38. Синтез гемоглобина Гем индуцируется синтез цепей глобина на рибосомах. Гены цепей глобина расположены в 11 и
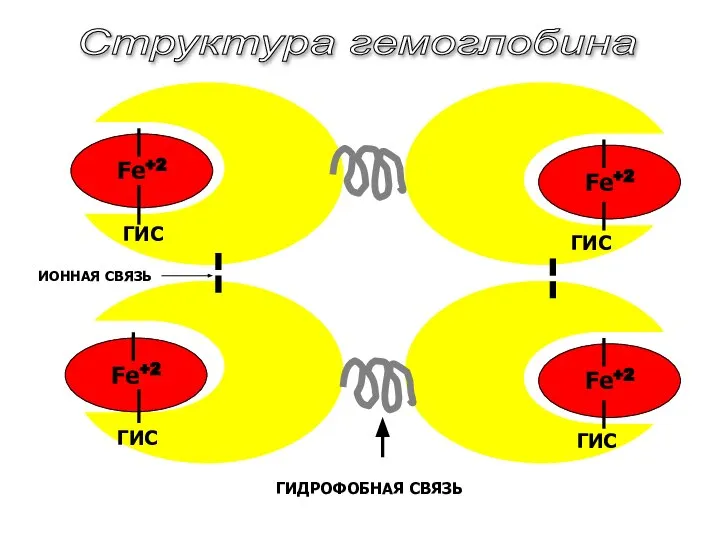
- 39. Fe+2 Fe+2 Fe+2 Fe+2 ГИДРОФОБНАЯ СВЯЗЬ ИОННАЯ СВЯЗЬ Структура гемоглобина ГИС ГИС ГИС ГИС
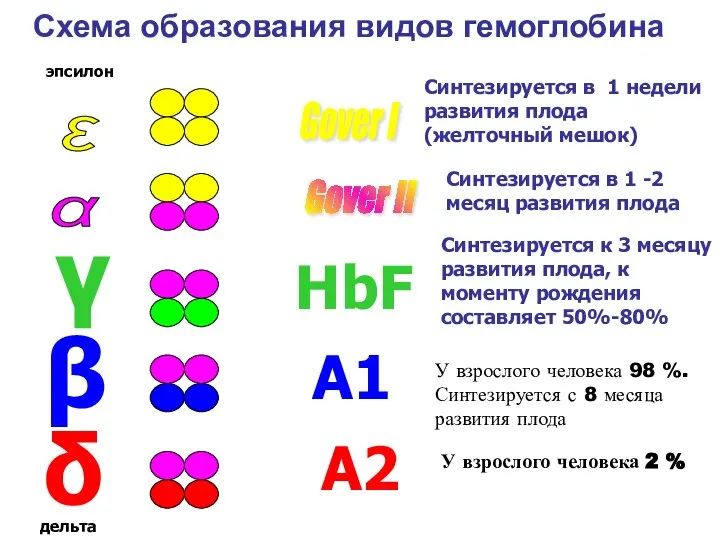
- 40. Виды гемоглобина Физиологические Эмбриональный: Gover I -4ε, Gover II -2α 2ε Фетальный - 2 α 2γ
- 41. ε α Gover I Gover II У взрослого человека 98 %. Синтезируется с 8 месяца развития
- 42. Талассемия Генетическое заболевание, обусловленное отсутствием или снижением синтеза одной из цепей гемоглобина. Причиной являются мутации генов-операторов,
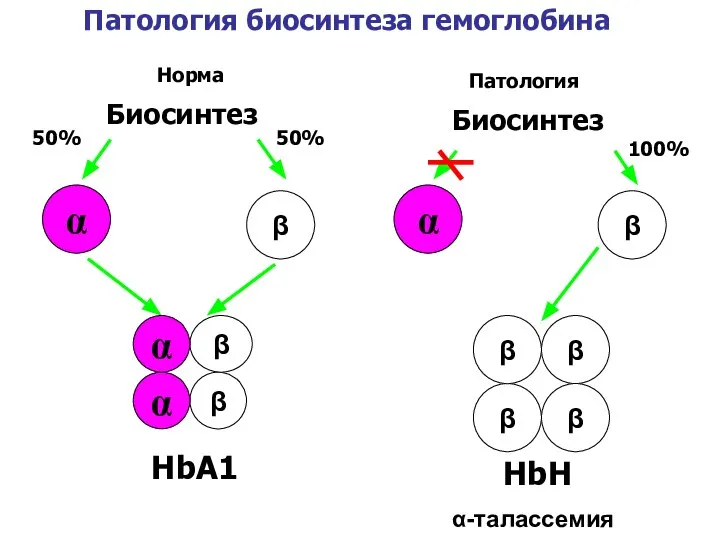
- 43. HbА1 Биосинтез α β 50% 50% α-талассемия Биосинтез α β 100% β β β β β
- 44. β-Талассемия Симптомы через полгода после рождения, когда прекращается синтез γ-цепи НвF. Прогрессирует анемия. Увеличиваются селезенка и
- 45. Серповидноклеточная анемия β-субъединицах НвS в шестом положении вместо ГЛУ находиться ВАЛ. Гидрофобные участок НвS, соединяются с
- 46. Механизм развития СКА О2 ВАЛ О2
- 47. Fe+2 ГИС О2 CO CN- OH- ОКСИГЕМОГЛОБИН КАРБОКСИГЕМОГЛОБИН МЕТГЕМОГЛОБИН ЦИАНМЕТГЕМОГЛОБИН Производные гемоглобина +3 ẽ ДЕЗОКСИГЕМОГЛОБИН
- 48. оксигемоглобин HbО2 (Fe2+) - соединение молекулярного кислорода с гемоглобином. Процесс называется оксигенацией, обратный процесс - дезоксигенацией
- 49. Производные гемоглобина: Карбоксигемоглобин HbСО (Fe2+) Связь гема с СО в двести раз прочнее, чем с О2.
- 50. метгемоглобин МtHb (Fe3+) Образуется при воздействии на гемоглобин окислителей (оксидов азота, хлоратов). В норме в крови
- 51. Цианметгемоглобин HbСN (Fe3+) образуется при присоединении СN- к метгемоглобину. Эта реакция спасает организм от смертельного действия
- 52. Fe+2 NH2 CO2 ОБРАЗОВАНИЕ КАРБГЕМОГЛОБИНА + ГИС NH-СООН
- 54. Аллостерическая регуляция насыщения гемоглобина кислородом На насыщение гемоглобина О2 влияют: Температура рH – эффект Бора Концентрация
- 55. Кривая диссоциации О2 для Hb ↑t◦ pH pCO2 2,3-ДФК
- 56. Fe+2 Fe+2 О2 Fe+2 Fe+2 гис гис гис гис О2 О2 О2 2,3-ФГК Сердце Головной мозг
- 57. Обмен веществ в лейкоцитах - полноценные клетки, содержат все органеллы. - присущи все виды обмена (кроме
- 59. АФК АФК используются фагоцитами для разрушения вирусов, онкоклеток оксидаза НАДФН2 + 2О2 -------------> НАДФН+ + 2О2²-
- 60. Образование NO L-АРГИНИН =(NO-СИНТАЗА)= L-ЦИТРУЛЛИН НАДФН НАДФ+ О2 NO
- 61. ФАГОЦИТОЗ Фагоцитоз-процесс активного поглощения и переваривания клетками организма попавших в него живых и убитых микробов или
- 62. Этапы фагоцитоза
- 66. Скачать презентацию

План лекции
Эритроцит: особенности структуры, химического состава, метаболизма белков, жиров, углеводов.
Гемоглобин и
План лекции
Эритроцит: особенности структуры, химического состава, метаболизма белков, жиров, углеводов.
Гемоглобин и

Эритроциты
форменные элементы крови
образуются в костном мозге,
циркулируют в крови около 100-120
Эритроциты
форменные элементы крови
образуются в костном мозге,
циркулируют в крови около 100-120
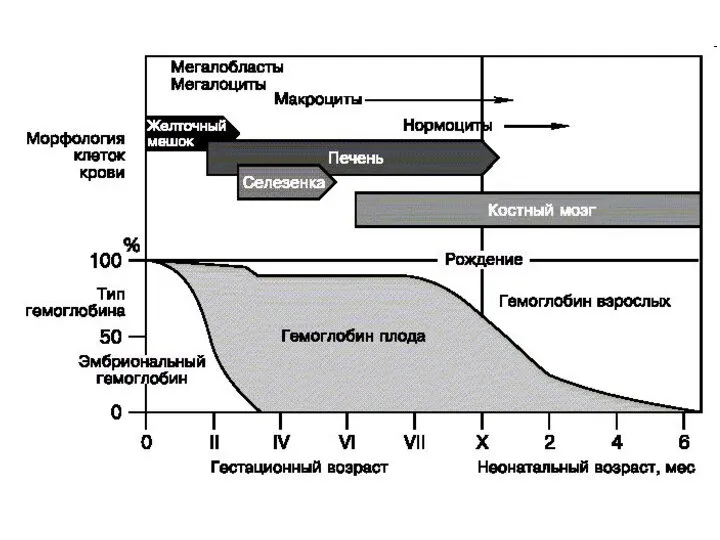

ФУНКЦИИ ЭРИТРОЦИТОВ
Газообмен: транспорт О2 и СО2
Регуляция КОС
Адсорбция и транспорт на клеточной
ФУНКЦИИ ЭРИТРОЦИТОВ
Газообмен: транспорт О2 и СО2
Регуляция КОС
Адсорбция и транспорт на клеточной

Строение эритроцита
60 % вода
40% сухой остаток
5 % - органические
и неорганические
Строение эритроцита
60 % вода
40% сухой остаток
5 % - органические
и неорганические
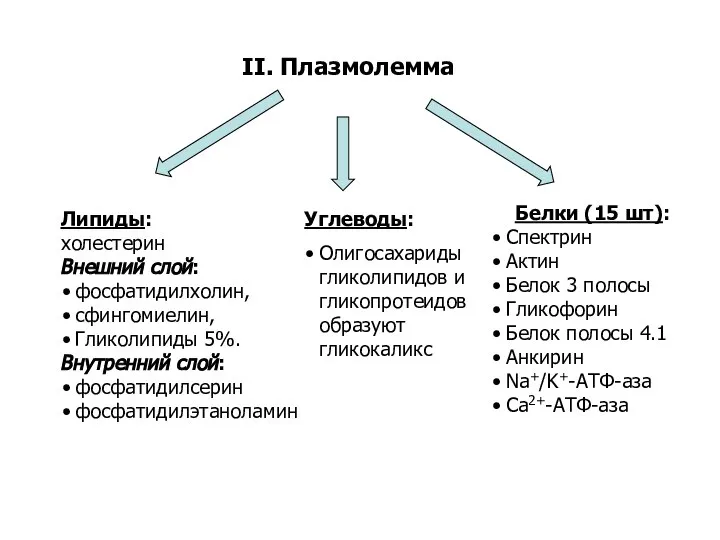
II. Плазмолемма
Белки (15 шт):
Спектрин
Актин
Белок 3 полосы
Гликофорин
Белок полосы 4.1
Анкирин
Na+/K+-АТФ-аза
Са2+-АТФ-аза
II. Плазмолемма
Белки (15 шт):
Спектрин
Актин
Белок 3 полосы
Гликофорин
Белок полосы 4.1
Анкирин
Na+/K+-АТФ-аза
Са2+-АТФ-аза

Часть белков плазмолеммы
образуют цитоскелет
фибриллярный белок спектрин образует сеть
глобулярные белки, пронизывающие
Часть белков плазмолеммы
образуют цитоскелет
фибриллярный белок спектрин образует сеть
глобулярные белки, пронизывающие
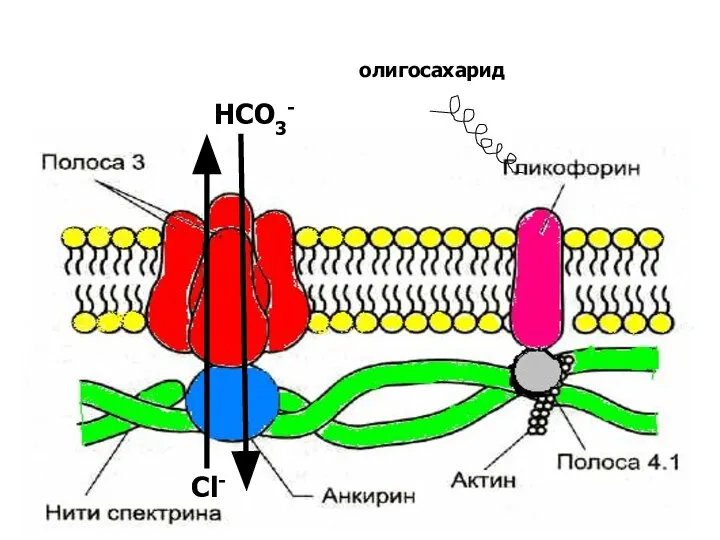
олигосахарид
Cl-
HCO3-
олигосахарид
Cl-
HCO3-


Спектрин
основной белок цитоскелета эритроцитов ( 25% массы всех белков),
имеет
Спектрин
основной белок цитоскелета эритроцитов ( 25% массы всех белков),
имеет

Гликофорин
трансмембранный гликопротеин (30 кДа), пронизывает плазмолемму в виде спирали.
Гликофорин
трансмембранный гликопротеин (30 кДа), пронизывает плазмолемму в виде спирали.

Углеводы
олигосахариды гликолипидов и гликопротеидов, на внешней поверхности плазмолеммы образуют гликокаликс.
Углеводы
олигосахариды гликолипидов и гликопротеидов, на внешней поверхности плазмолеммы образуют гликокаликс.

ОСОБЕННОСТЬ ОБМЕНА ВЕЩЕСТВ
И ЭНЕРГИИ В ЭРИТРОЦИТЕ
Эритроцит высокоспециализированная клетка, хорошо приспособленная
ОСОБЕННОСТЬ ОБМЕНА ВЕЩЕСТВ
И ЭНЕРГИИ В ЭРИТРОЦИТЕ
Эритроцит высокоспециализированная клетка, хорошо приспособленная

Особенности обмена нуклеотидов
рибозо-5ф + аденин → АМФ
АМФ + АТФ
Особенности обмена нуклеотидов
рибозо-5ф + аденин → АМФ
АМФ + АТФ
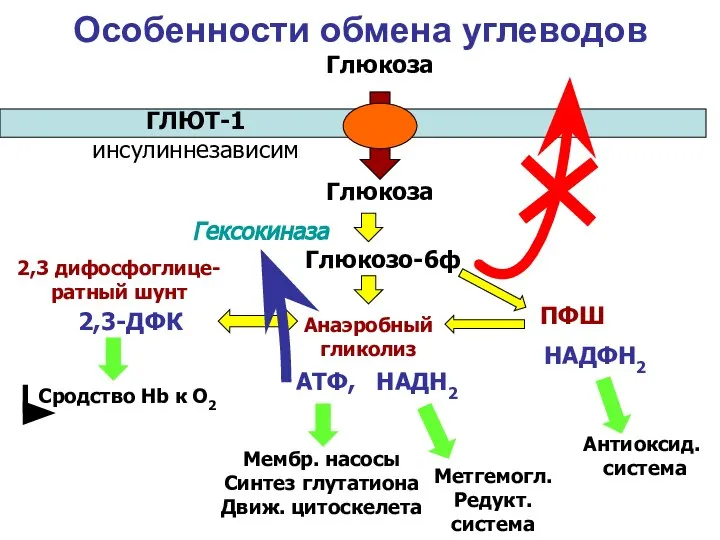
Особенности обмена углеводов
Анаэробный
гликолиз
ПФШ
2,3 дифосфоглице-
ратный шунт
Глюкоза
Глюкоза
Глюкозо-6ф
ГЛЮТ-1 инсулиннезависим
Гексокиназа
АТФ, НАДН2
НАДФН2
2,3-ДФК
Мембр. насосы
Синтез глутатиона
Движ.
Особенности обмена углеводов
Анаэробный
гликолиз
ПФШ
2,3 дифосфоглице-
ратный шунт
Глюкоза
Глюкоза
Глюкозо-6ф
ГЛЮТ-1 инсулиннезависим
Гексокиназа
АТФ, НАДН2
НАДФН2
2,3-ДФК
Мембр. насосы
Синтез глутатиона
Движ.

(метод оценки ПРЭ)
(метод оценки ПРЭ)

Обмен метгемоглобина
В течение суток до 3% гемоглобина может спонтанно окисляться в
Обмен метгемоглобина
В течение суток до 3% гемоглобина может спонтанно окисляться в
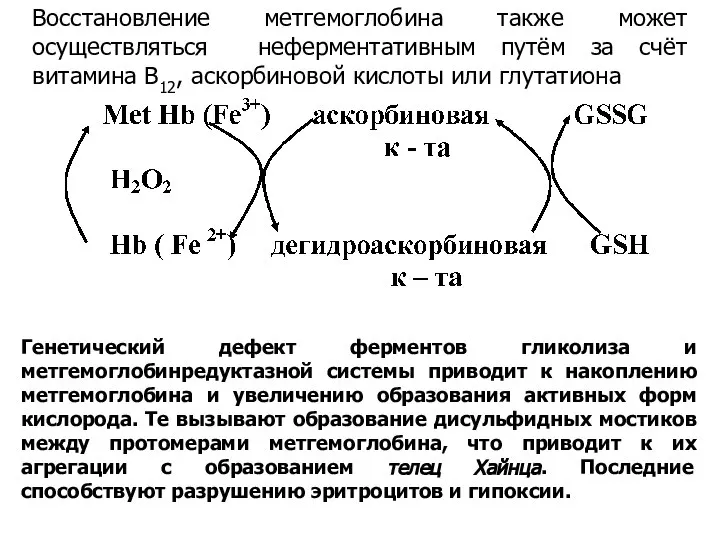
Восстановление метгемоглобина также может осуществляться неферментативным путём за счёт витамина В12,
Восстановление метгемоглобина также может осуществляться неферментативным путём за счёт витамина В12,
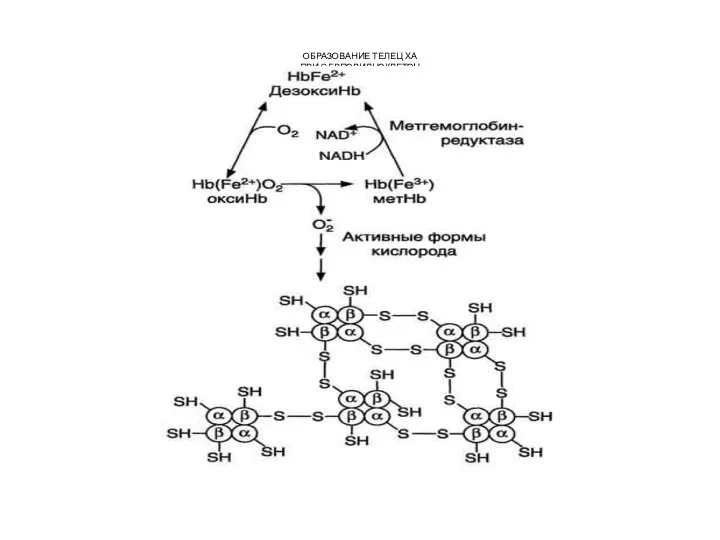
ОБРАЗОВАНИЕ ТЕЛЕЦ ХА
ПРИ СЕРПОВИДНОКЛЕТОЧ
НОЙ АНЕМИИ
ОБРАЗОВАНИЕ ТЕЛЕЦ ХА
ПРИ СЕРПОВИДНОКЛЕТОЧ
НОЙ АНЕМИИ

Na+/K+-АТФ-аза - мембранный фермент поддерживает градиент концентраци Na+ и К+
Патология
Na+/K+-АТФ-аза - мембранный фермент поддерживает градиент концентраци Na+ и К+
Патология


Особенности липидного обмена
В зрелом эритроците липиды не синтезируются, эритроцит
Особенности липидного обмена
В зрелом эритроците липиды не синтезируются, эритроцит

В зрелом эритроците белки не синтезируются, т.к. у него нет
В зрелом эритроците белки не синтезируются, т.к. у него нет
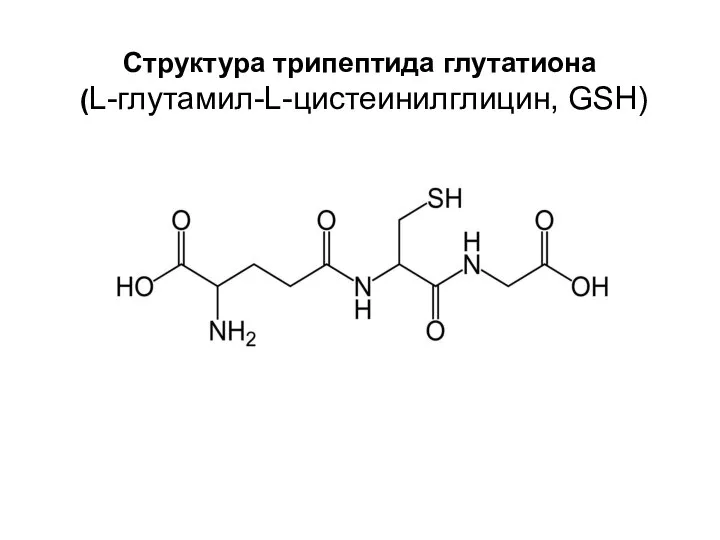
Структура трипептида глутатиона
(L-глутамил-L-цистеинилглицин, GSH)
Структура трипептида глутатиона
(L-глутамил-L-цистеинилглицин, GSH)

Биосинтез глутатиона
Глутатион синтезируется из аминокислот L-цистеина, L-глутаминовой кислоты и глицина.
Синтез происходит в две АТР-зависимые стадии:
1.
Биосинтез глутатиона
Глутатион синтезируется из аминокислот L-цистеина, L-глутаминовой кислоты и глицина.
Синтез происходит в две АТР-зависимые стадии:
1.

Особенность метаболизма при старении эритроцитов
↓ образование АТФ
Нарушаются энергозависимые процессы
Особенность метаболизма при старении эритроцитов
↓ образование АТФ
Нарушаются энергозависимые процессы
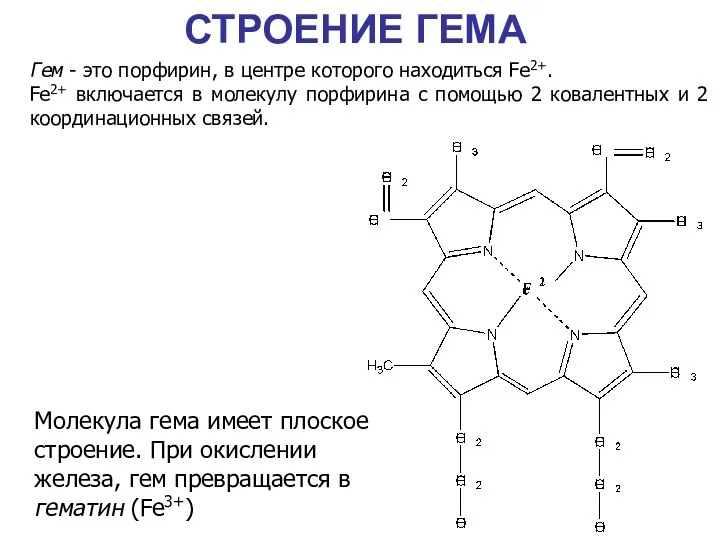
СТРОЕНИЕ ГЕМА
Гем - это порфирин, в центре которого находиться Fe2+.
Fe2+ включается
СТРОЕНИЕ ГЕМА
Гем - это порфирин, в центре которого находиться Fe2+.
Fe2+ включается

Наибольшее количество гема содержат эритроциты (Hb), мышечные клетки (миоглобин) и клетки
Наибольшее количество гема содержат эритроциты (Hb), мышечные клетки (миоглобин) и клетки
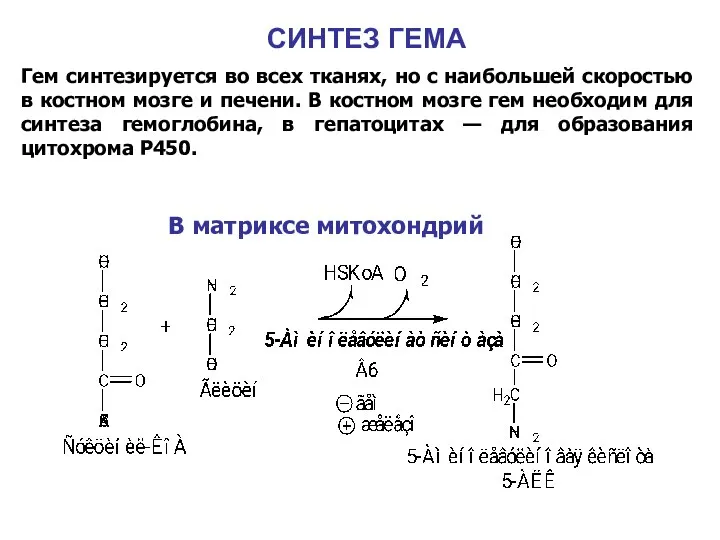
СИНТЕЗ ГЕМА
Гем синтезируется во всех тканях, но с наибольшей скоростью в
СИНТЕЗ ГЕМА
Гем синтезируется во всех тканях, но с наибольшей скоростью в
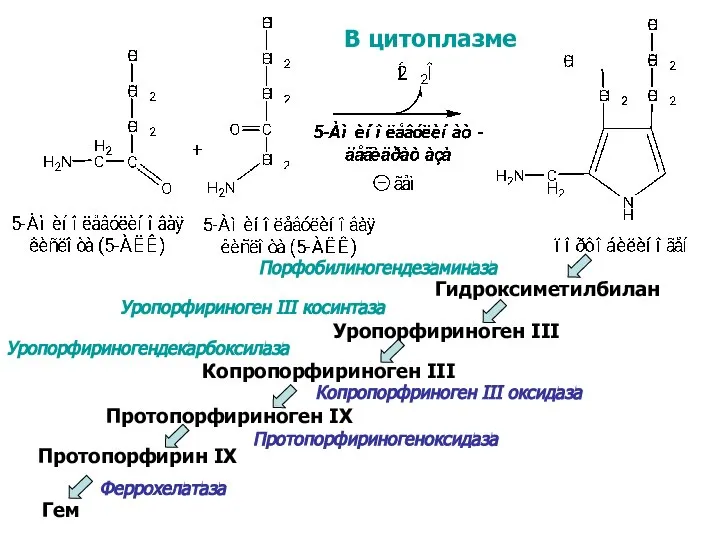
В цитоплазме
Гидроксиметилбилан
Уропорфириноген III
Копропорфириноген III
Протопорфириноген IX
Протопорфирин IX
Гем
Порфобилиногендезаминаза
Уропорфириноген
В цитоплазме
Гидроксиметилбилан
Уропорфириноген III
Копропорфириноген III
Протопорфириноген IX
Протопорфирин IX
Гем
Порфобилиногендезаминаза
Уропорфириноген

Порфирии - гетерогенная группа заболеваний, вызванная нарушениями синтеза гема вследствие дефицита
Порфирии - гетерогенная группа заболеваний, вызванная нарушениями синтеза гема вследствие дефицита


Негативные последствия порфирий связаны с дефицитом гема и накоплением в организме
Негативные последствия порфирий связаны с дефицитом гема и накоплением в организме
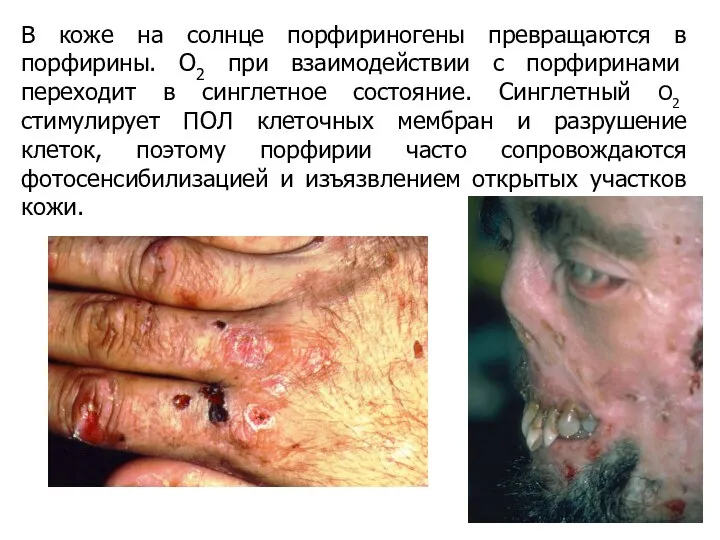
В коже на солнце порфириногены превращаются в порфирины. О2 при взаимодействии
В коже на солнце порфириногены превращаются в порфирины. О2 при взаимодействии

Порфириногены бесцветны и не флуоресцируют, а порфирины имеют интенсивную красную флуоресценцию
Порфириногены бесцветны и не флуоресцируют, а порфирины имеют интенсивную красную флуоресценцию
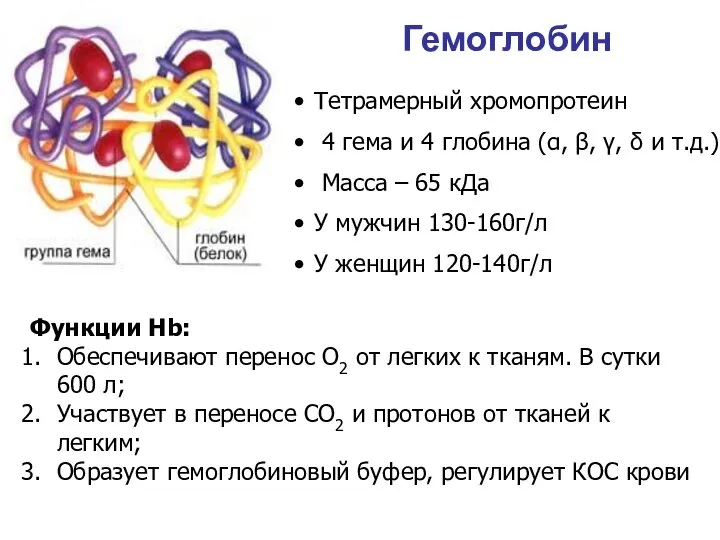
Гемоглобин
Тетрамерный хромопротеин
4 гема и 4 глобина (α, β, γ, δ
Гемоглобин
Тетрамерный хромопротеин
4 гема и 4 глобина (α, β, γ, δ

Синтез гемоглобина
Гем индуцируется синтез цепей глобина на рибосомах. Гены цепей глобина
Синтез гемоглобина
Гем индуцируется синтез цепей глобина на рибосомах. Гены цепей глобина
Fe+2
Fe+2
Fe+2
Fe+2
ГИДРОФОБНАЯ СВЯЗЬ
ИОННАЯ СВЯЗЬ
Структура гемоглобина
ГИС
ГИС
ГИС
ГИС
Fe+2
Fe+2
Fe+2
Fe+2
ГИДРОФОБНАЯ СВЯЗЬ
ИОННАЯ СВЯЗЬ
Структура гемоглобина
ГИС
ГИС
ГИС
ГИС

Виды гемоглобина
Физиологические
Эмбриональный:
Gover I -4ε,
Gover II -2α 2ε
Фетальный - 2
Виды гемоглобина
Физиологические
Эмбриональный:
Gover I -4ε,
Gover II -2α 2ε
Фетальный - 2
ε
α
Gover I
Gover II
У взрослого человека 98 %.
ε
α
Gover I
Gover II
У взрослого человека 98 %.

Талассемия
Генетическое заболевание, обусловленное отсутствием или снижением синтеза одной из цепей
Талассемия
Генетическое заболевание, обусловленное отсутствием или снижением синтеза одной из цепей
HbА1
Биосинтез
α
β
50%
50%
α-талассемия
Биосинтез
α
β
100%
β
β
β
β
β
β
α
α
Патология биосинтеза гемоглобина
Патология
Норма
HbН
HbА1
Биосинтез
α
β
50%
50%
α-талассемия
Биосинтез
α
β
100%
β
β
β
β
β
β
α
α
Патология биосинтеза гемоглобина
Патология
Норма
HbН
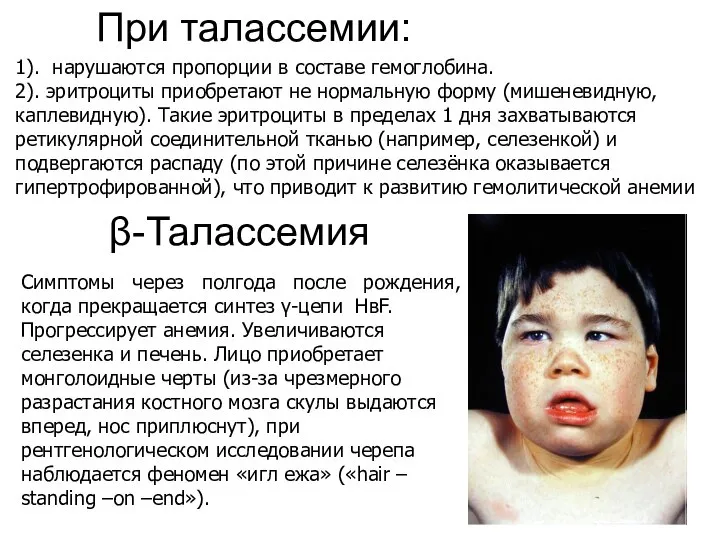
β-Талассемия
Симптомы через полгода после рождения, когда прекращается синтез γ-цепи НвF.
Прогрессирует
β-Талассемия
Симптомы через полгода после рождения, когда прекращается синтез γ-цепи НвF.
Прогрессирует

Серповидноклеточная анемия
β-субъединицах НвS в шестом положении вместо ГЛУ находиться ВАЛ.
Серповидноклеточная анемия
β-субъединицах НвS в шестом положении вместо ГЛУ находиться ВАЛ.
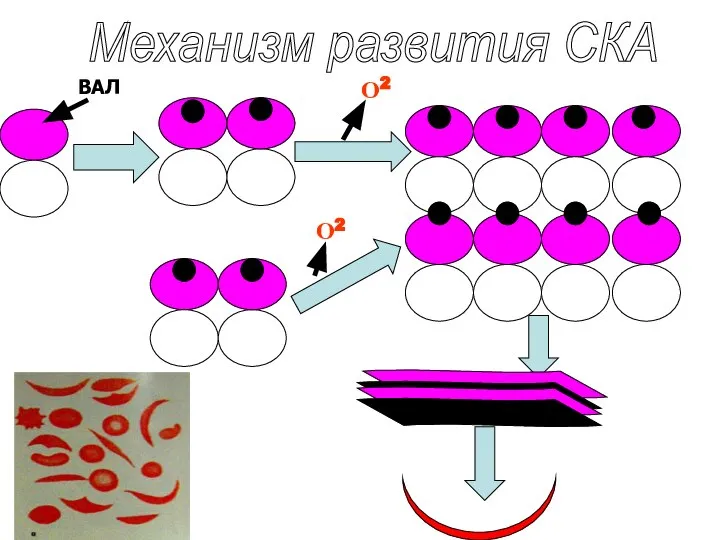
Механизм развития СКА
О2
ВАЛ
О2
Механизм развития СКА
О2
ВАЛ
О2
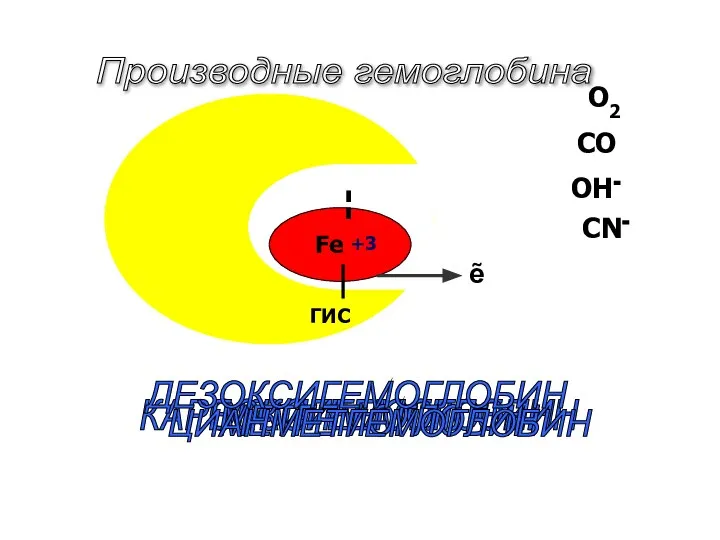
Fe+2
ГИС
О2
CO
CN-
OH-
ОКСИГЕМОГЛОБИН
КАРБОКСИГЕМОГЛОБИН
МЕТГЕМОГЛОБИН
ЦИАНМЕТГЕМОГЛОБИН
Производные гемоглобина
+3
ẽ
ДЕЗОКСИГЕМОГЛОБИН
Fe+2
ГИС
О2
CO
CN-
OH-
ОКСИГЕМОГЛОБИН
КАРБОКСИГЕМОГЛОБИН
МЕТГЕМОГЛОБИН
ЦИАНМЕТГЕМОГЛОБИН
Производные гемоглобина
+3
ẽ
ДЕЗОКСИГЕМОГЛОБИН

оксигемоглобин HbО2 (Fe2+)
- соединение молекулярного кислорода с гемоглобином.
Процесс называется
оксигемоглобин HbО2 (Fe2+)
- соединение молекулярного кислорода с гемоглобином.
Процесс называется

Производные гемоглобина:
Карбоксигемоглобин HbСО (Fe2+)
Связь гема с СО
Производные гемоглобина:
Карбоксигемоглобин HbСО (Fe2+)
Связь гема с СО

метгемоглобин МtHb (Fe3+)
Образуется при воздействии на гемоглобин окислителей (оксидов азота, хлоратов).
метгемоглобин МtHb (Fe3+)
Образуется при воздействии на гемоглобин окислителей (оксидов азота, хлоратов).

Цианметгемоглобин HbСN (Fe3+)
образуется при присоединении СN- к метгемоглобину.
Цианметгемоглобин HbСN (Fe3+)
образуется при присоединении СN- к метгемоглобину.
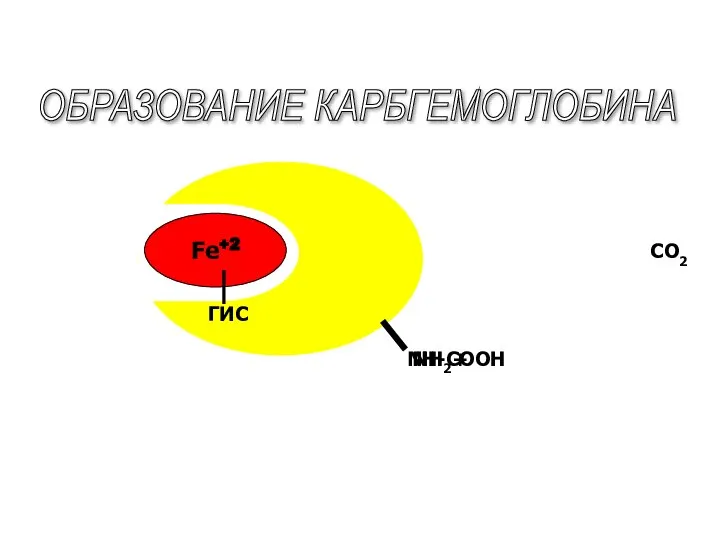
Fe+2
NH2
CO2
ОБРАЗОВАНИЕ КАРБГЕМОГЛОБИНА
+
ГИС
NH-СООН
Fe+2
NH2
CO2
ОБРАЗОВАНИЕ КАРБГЕМОГЛОБИНА
+
ГИС
NH-СООН

Аллостерическая регуляция насыщения гемоглобина кислородом
На насыщение гемоглобина О2 влияют:
Температура
рH – эффект
Аллостерическая регуляция насыщения гемоглобина кислородом
На насыщение гемоглобина О2 влияют:
Температура
рH – эффект
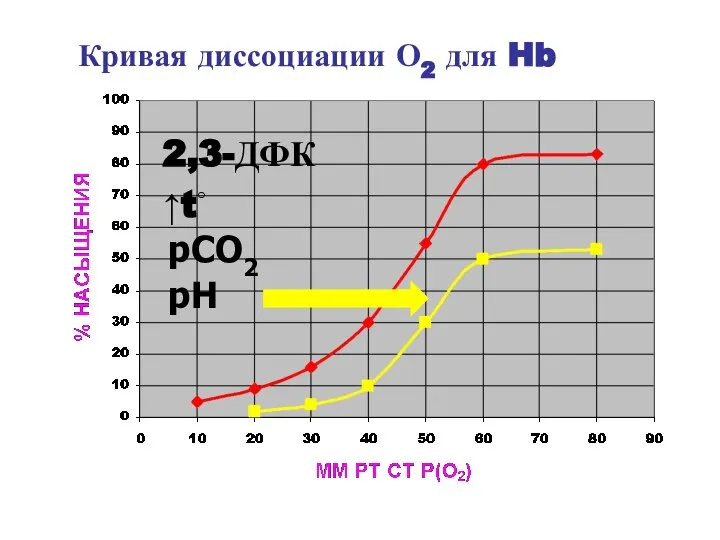
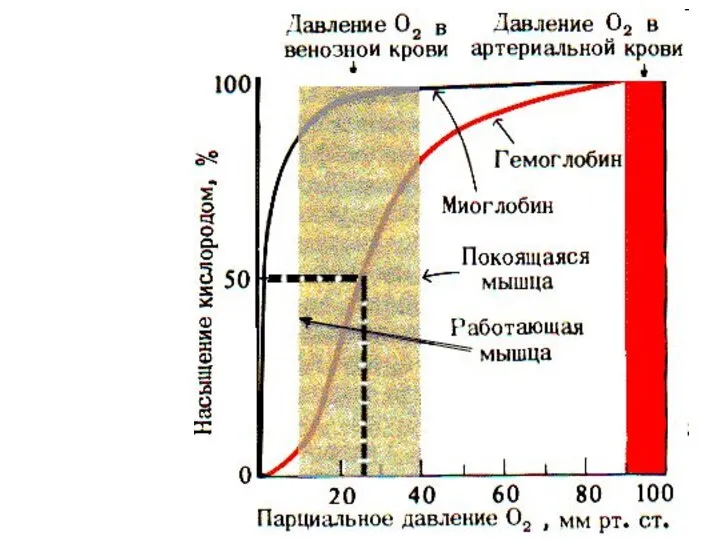
Кривая диссоциации О2 для Hb
↑t◦
pH
pCO2
2,3-ДФК
Кривая диссоциации О2 для Hb
↑t◦
pH
pCO2
2,3-ДФК
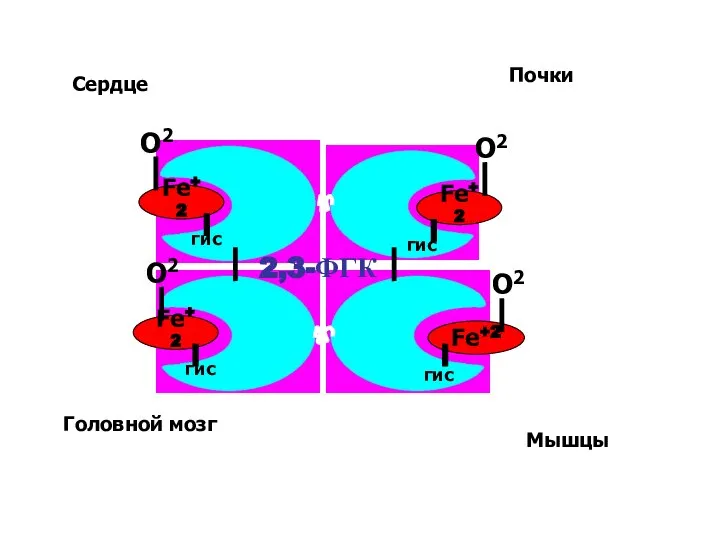
Fe+2
Fe+2
О2
Fe+2
Fe+2
гис
гис
гис
гис
О2
О2
О2
2,3-ФГК
Сердце
Головной мозг
Почки
Мышцы
Fe+2
Fe+2
О2
Fe+2
Fe+2
гис
гис
гис
гис
О2
О2
О2
2,3-ФГК
Сердце
Головной мозг
Почки
Мышцы

Обмен веществ в лейкоцитах
- полноценные клетки, содержат все органеллы.
- присущи все
Обмен веществ в лейкоцитах
- полноценные клетки, содержат все органеллы.
- присущи все

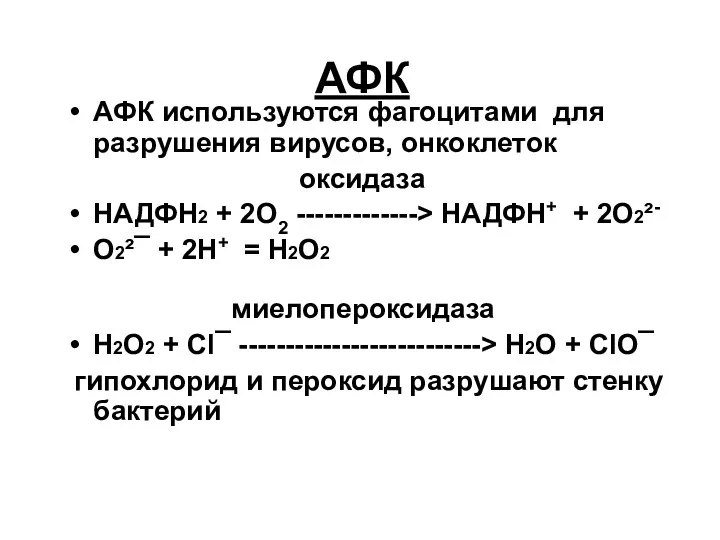
АФК
АФК используются фагоцитами для разрушения вирусов, онкоклеток
оксидаза
НАДФН2 + 2О2
АФК
АФК используются фагоцитами для разрушения вирусов, онкоклеток
оксидаза
НАДФН2 + 2О2
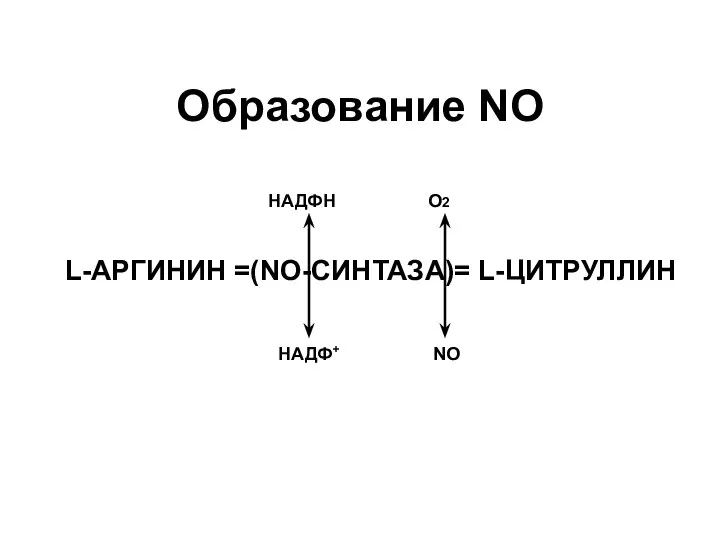
Образование NO
L-АРГИНИН =(NO-СИНТАЗА)= L-ЦИТРУЛЛИН
НАДФН
НАДФ+
О2
NO
Образование NO
L-АРГИНИН =(NO-СИНТАЗА)= L-ЦИТРУЛЛИН
НАДФН
НАДФ+
О2
NO

ФАГОЦИТОЗ
Фагоцитоз-процесс активного поглощения и переваривания клетками организма попавших в него живых
ФАГОЦИТОЗ
Фагоцитоз-процесс активного поглощения и переваривания клетками организма попавших в него живых
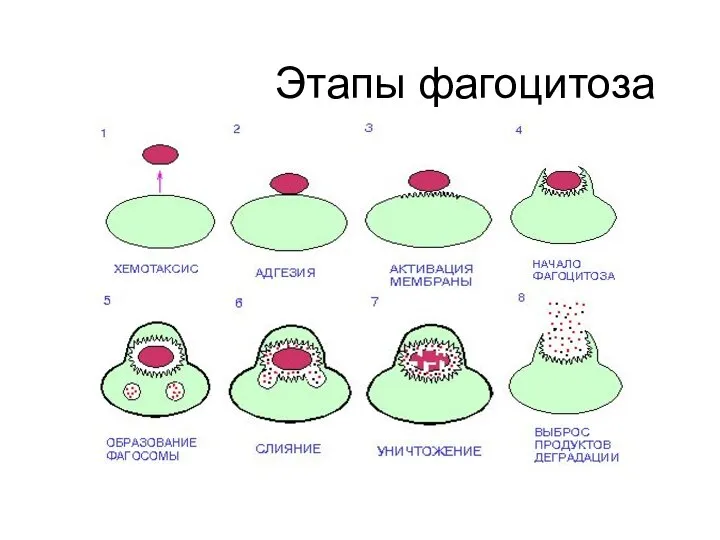
Этапы фагоцитоза
Этапы фагоцитоза


 Химический элемент - америций
Химический элемент - америций Введение. Теория строения органических соединений А.М.Бутлерова. Лекция № 1
Введение. Теория строения органических соединений А.М.Бутлерова. Лекция № 1 Химический состав клетки
Химический состав клетки Волокнистые наполнители. (Тема 5)
Волокнистые наполнители. (Тема 5) Классификация неорганических соединений. Оксиды и их свойства
Классификация неорганических соединений. Оксиды и их свойства Гормоны
Гормоны  Алифатические углеводороды. Алканы, их свойства
Алифатические углеводороды. Алканы, их свойства Лекция 9-10. Диффузионная кинетика
Лекция 9-10. Диффузионная кинетика Залежність властивостей полімерів від їхньої будови. Термопластичні й термореактивні полімери
Залежність властивостей полімерів від їхньої будови. Термопластичні й термореактивні полімери Химический элемент. Вещества
Химический элемент. Вещества Окислительно-восстановительные реакции. (лекция №10)
Окислительно-восстановительные реакции. (лекция №10) Насыщенный пар. Зависимость давления насыщенного пара от температуры. Кипение
Насыщенный пар. Зависимость давления насыщенного пара от температуры. Кипение Фуллерены (шары Бакминстера)
Фуллерены (шары Бакминстера) Аппаратура ионообменных процессов
Аппаратура ионообменных процессов Игровые моменты на уроках химии Подготовила: Трубчанинова Е.П., учитель биологии и химии Грабской ЗОШ І-ІІ ст. Амвросиевского р
Игровые моменты на уроках химии Подготовила: Трубчанинова Е.П., учитель биологии и химии Грабской ЗОШ І-ІІ ст. Амвросиевского р Охорона навколишнього середовища від забруднення викидами сульфуровмісних речовин.
Охорона навколишнього середовища від забруднення викидами сульфуровмісних речовин.  Спирт. Спирты в природе. Влияние спирта на человека
Спирт. Спирты в природе. Влияние спирта на человека Неметаллы. Общая характеристика
Неметаллы. Общая характеристика Природный и попутный нефтяной газ
Природный и попутный нефтяной газ Підгрупа карбону
Підгрупа карбону Электронные эффекты заместителей. Типы химических реакций
Электронные эффекты заместителей. Типы химических реакций Роль химии в нашей жизни
Роль химии в нашей жизни Защита полимеров от старения
Защита полимеров от старения Сплавы
Сплавы Технологія жирів і жирозамінників
Технологія жирів і жирозамінників Электродные процессы, их биологическая роль и применение в медицине
Электродные процессы, их биологическая роль и применение в медицине Нанокаталіз. Активність каталізатора
Нанокаталіз. Активність каталізатора Классификация опасных грузов
Классификация опасных грузов