- Строение и функции нуклеиновых кислот

Содержание
- 2. Нуклеиновые кислоты (полинуклеотиды) - это биополимеры, мономерными звеньями которых являются нуклеотиды.
- 3. Нуклеиновые основания Пиримидиновые основания в составе полинуклеотидов представлены лактамной формой, что обусловлено возможностью образования водородных связей
- 4. Нуклеиновые основания Аденин и гуанин являются представителями пуриновых нуклеиновых оснований. Эти гетероциклические соединения способны к лактим-лактамной
- 5. Редкие (минорные) компоненты нуклеиновых кислот В ДНК встречаются метилированные основания: 5-метилцитозин, 6-N- метиладенин и др. В
- 6. Нуклеозиды Гетероциклические основания образуют N-гликозиды (нуклеозиды) с D-рибозой или 2-дезокси-D-рибозой.. D-рибоза и 2-дезокси-D-рибоза в состав природных
- 7. Структура нуклеозидов
- 8. В состав некоторых РНК входят необычные нуклеозиды. Например, инозин, который можно рассматривать как продукт дезаминирования аденозина,
- 9. Нуклеотиды Нуклеотиды - фосфаты нуклеозидов. Фосфорная кислота обычно этерифицирует спиртовый гидроксил при С-5' или С-3' в
- 10. Структура некоторых нуклеотидов
- 11. Макроэргические связи — ковалентные связи, которые гидролизуются с выделением значительного количества энергии: 30 кДж/моль и более.
- 12. Строение полинуклеотидов Многообразие молекул ДНК и РНК определяется их первичной структурой – последовательностью нуклеотидных остатков в
- 13. Первичная структура нуклеиновых кислот Первичная структура нуклеиновых кислот – нуклеотидный состав и нуклеотидная последовательность, т.е. порядок
- 14. Вторичная структура ДНК Вторичная структура ДНК – это пространственная организация полинуклеотидной цепи. Водородные связи образуются между
- 15. Молекула РНК построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли – «шпильки»,
- 17. Скачать презентацию
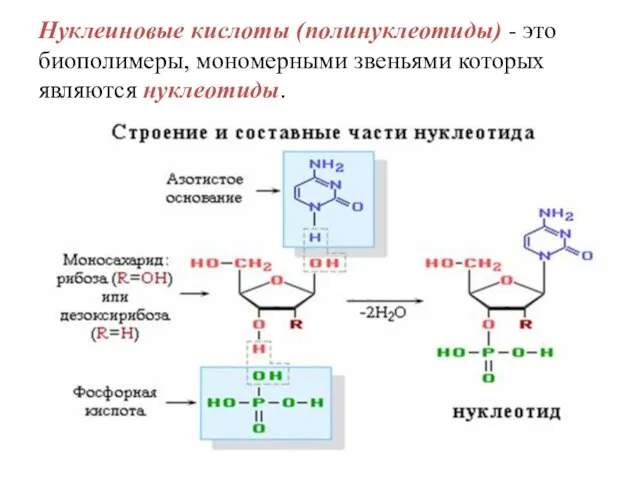
Нуклеиновые кислоты (полинуклеотиды) - это биополимеры, мономерными звеньями которых являются нуклеотиды.
Нуклеиновые кислоты (полинуклеотиды) - это биополимеры, мономерными звеньями которых являются нуклеотиды.
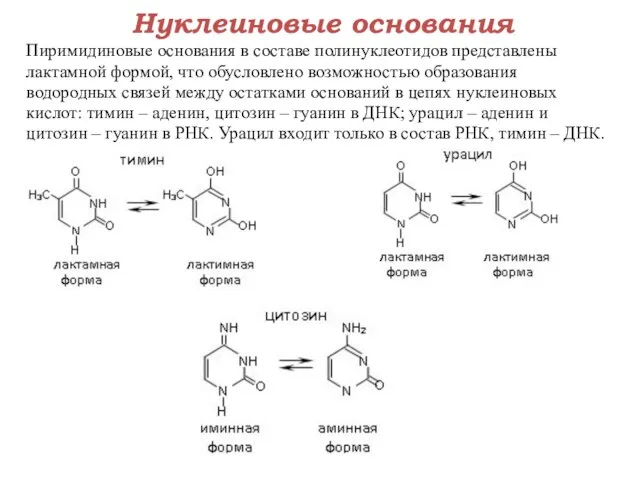
Нуклеиновые основания
Пиримидиновые основания в составе полинуклеотидов представлены
лактамной формой, что обусловлено
Нуклеиновые основания
Пиримидиновые основания в составе полинуклеотидов представлены
лактамной формой, что обусловлено
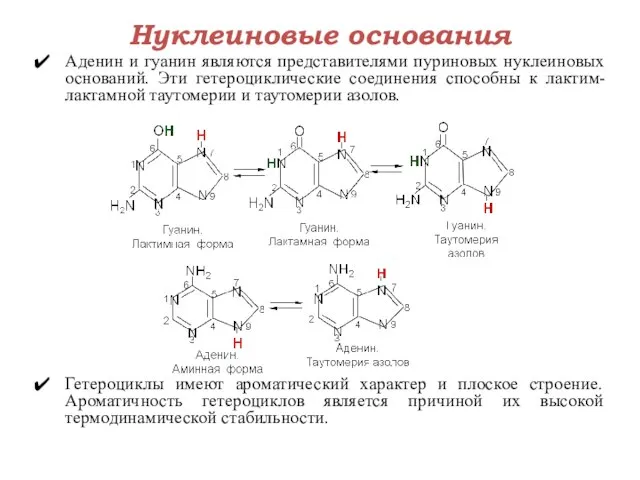
Нуклеиновые основания
Аденин и гуанин являются представителями пуриновых нуклеиновых оснований. Эти
Нуклеиновые основания
Аденин и гуанин являются представителями пуриновых нуклеиновых оснований. Эти
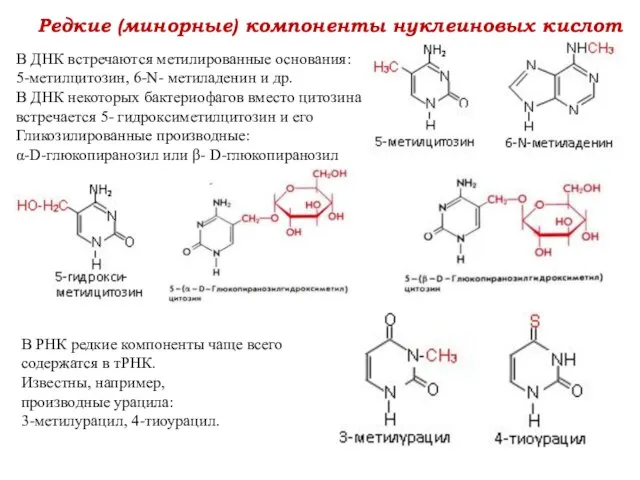
Редкие (минорные) компоненты нуклеиновых кислот
В ДНК встречаются метилированные основания:
5-метилцитозин, 6-N-
Редкие (минорные) компоненты нуклеиновых кислот
В ДНК встречаются метилированные основания:
5-метилцитозин, 6-N-

Нуклеозиды
Гетероциклические основания образуют N-гликозиды (нуклеозиды) с D-рибозой или 2-дезокси-D-рибозой..
D-рибоза и
Нуклеозиды
Гетероциклические основания образуют N-гликозиды (нуклеозиды) с D-рибозой или 2-дезокси-D-рибозой..
D-рибоза и
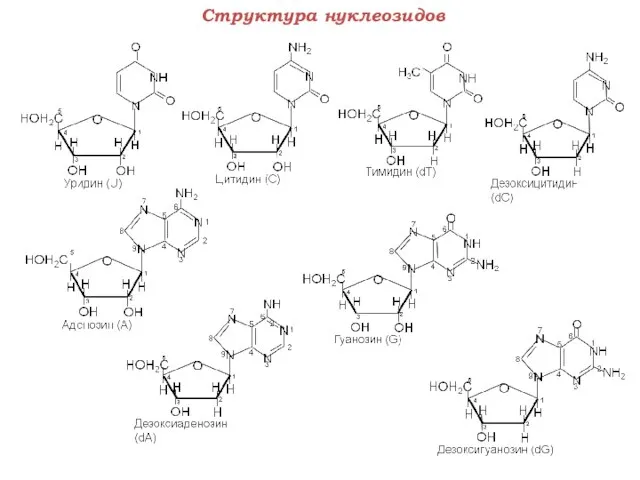
Структура нуклеозидов
Структура нуклеозидов
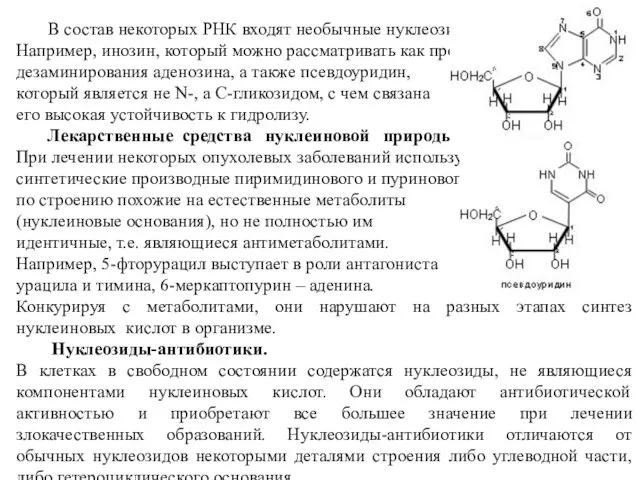
В состав некоторых РНК входят необычные нуклеозиды.
Например, инозин, который
В состав некоторых РНК входят необычные нуклеозиды.
Например, инозин, который

Нуклеотиды
Нуклеотиды - фосфаты нуклеозидов. Фосфорная кислота обычно этерифицирует спиртовый гидроксил
Нуклеотиды
Нуклеотиды - фосфаты нуклеозидов. Фосфорная кислота обычно этерифицирует спиртовый гидроксил
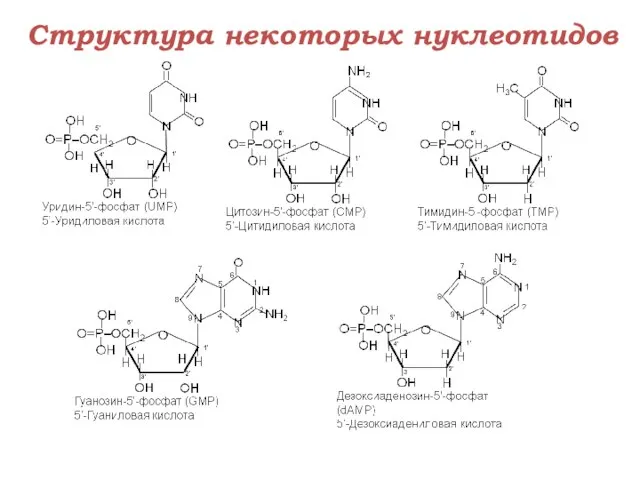
Структура некоторых нуклеотидов
Структура некоторых нуклеотидов
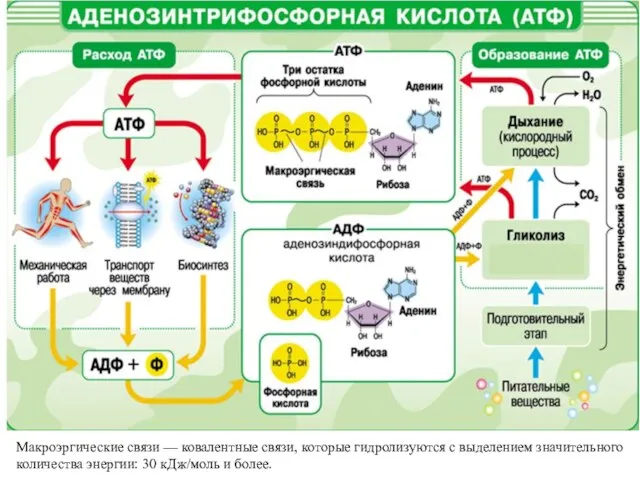
Макроэргические связи — ковалентные связи, которые гидролизуются с выделением значительного количества
Макроэргические связи — ковалентные связи, которые гидролизуются с выделением значительного количества
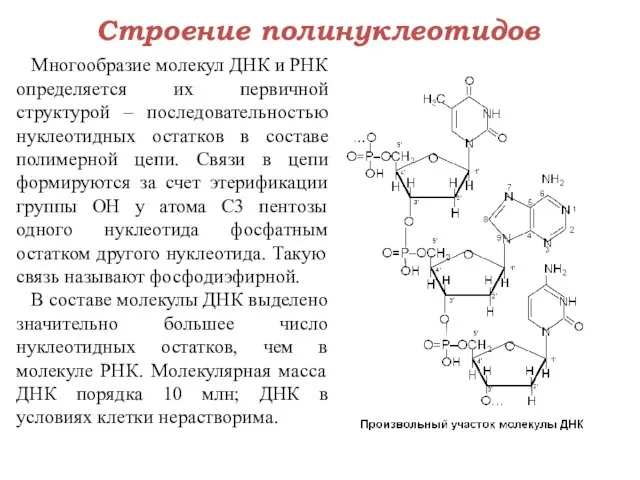
Строение полинуклеотидов
Многообразие молекул ДНК и РНК определяется их первичной структурой
Строение полинуклеотидов
Многообразие молекул ДНК и РНК определяется их первичной структурой

Первичная структура нуклеиновых кислот
Первичная структура нуклеиновых кислот – нуклеотидный состав и
Первичная структура нуклеиновых кислот
Первичная структура нуклеиновых кислот – нуклеотидный состав и
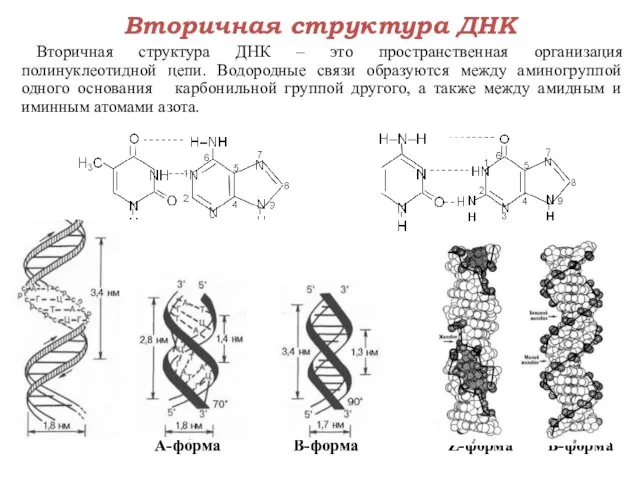
Вторичная структура ДНК
Вторичная структура ДНК – это пространственная организация полинуклеотидной цепи.
Вторичная структура ДНК
Вторичная структура ДНК – это пространственная организация полинуклеотидной цепи.
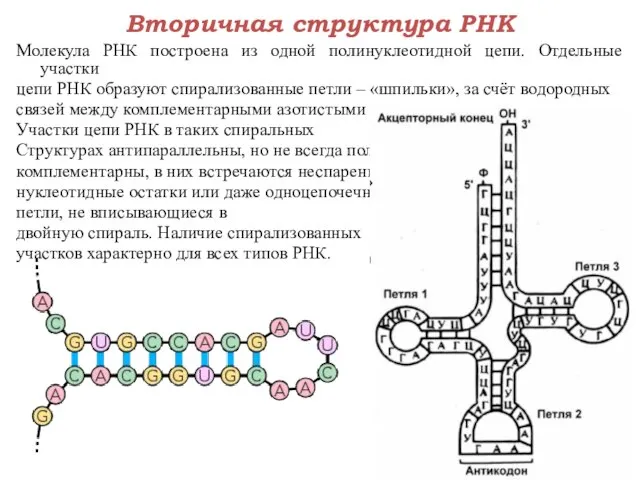
Молекула РНК построена из одной полинуклеотидной цепи. Отдельные участки
цепи РНК образуют
Молекула РНК построена из одной полинуклеотидной цепи. Отдельные участки
цепи РНК образуют
 Неметаллы. Смотр знаний. Для учащихся 9 классов
Неметаллы. Смотр знаний. Для учащихся 9 классов Сложные эфиры. Жиры
Сложные эфиры. Жиры Аттестационная работа. Методика по химии 9 класс Исследовательская деятельность в решении экспериментальных задач
Аттестационная работа. Методика по химии 9 класс Исследовательская деятельность в решении экспериментальных задач Насыщенные углеводороды. (Тема 1.1)
Насыщенные углеводороды. (Тема 1.1) Презентация по Химии "Оксид алюмінію" - скачать смотреть бесплатно
Презентация по Химии "Оксид алюмінію" - скачать смотреть бесплатно Занимательные опыты по химии
Занимательные опыты по химии Бүйрек зәр түзе отырып, қаннан шығатын бөлінділерді сүзетін
Бүйрек зәр түзе отырып, қаннан шығатын бөлінділерді сүзетін Займання та горіння твердих горючих матеріалів (лекція 11)
Займання та горіння твердих горючих матеріалів (лекція 11) Кислоты. Повторение
Кислоты. Повторение Эволюция понятия “валентность”
Эволюция понятия “валентность” Презентация Принципы гормональной и нейромедиаторной регуляции
Презентация Принципы гормональной и нейромедиаторной регуляции 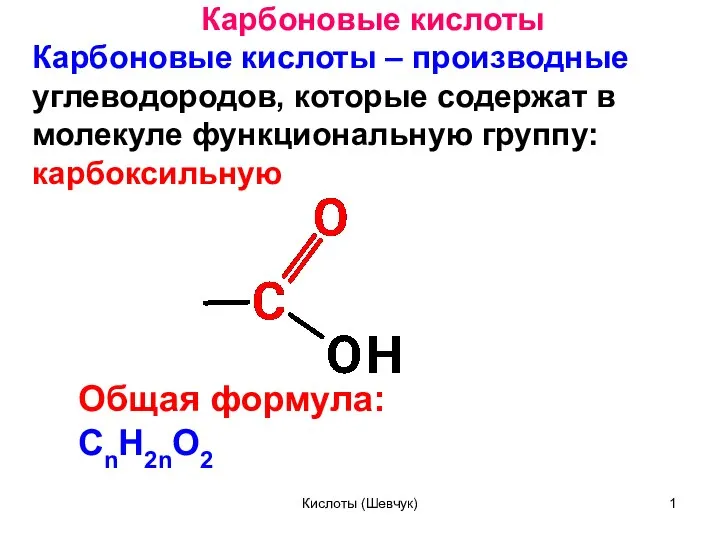 Карбоновые кислоты
Карбоновые кислоты Комплексные соединения
Комплексные соединения Значение минеральных веществ и витаминов в питании человека
Значение минеральных веществ и витаминов в питании человека Эндогенная серия. Скарновая группа
Эндогенная серия. Скарновая группа Ароматичні спирти
Ароматичні спирти Основы кристаллографии
Основы кристаллографии Происхождение рибосомы, белкового синтеза и генетического кода
Происхождение рибосомы, белкового синтеза и генетического кода Электролиз. Электролиз хлорида натрия
Электролиз. Электролиз хлорида натрия Термодинамика. Законы термодинамики
Термодинамика. Законы термодинамики Кристалы и их свойства
Кристалы и их свойства Амедео Авогадро
Амедео Авогадро Оксиды. Кварц (SiO2). Корунд (Al2O3)
Оксиды. Кварц (SiO2). Корунд (Al2O3) Основания, их классификация и свойства в свете ТЭД
Основания, их классификация и свойства в свете ТЭД Кварцевые пески
Кварцевые пески Химические волокна (7 класс)
Химические волокна (7 класс) Галогены. Галогеноводородные кислоты и их соли
Галогены. Галогеноводородные кислоты и их соли Кристаллохимические радиусы
Кристаллохимические радиусы