- Антитела. Основные формы проявления иммунитета

Содержание
- 2. Основные формы проявления иммунитета: антителообразование, иммунный фагоцитоз, опосредованный клетками киллинг, реакции гиперчувствительности, формирование иммунологической памяти, формирование
- 3. Антитела (иммуноглобулины) белки, специфически реагирующие с антигенами. Относятся к гамма-глобулиновой фракции белков сыворотки крови (15-25%, т.е.
- 4. Молекулярное строение антител 2 пары полипептидных цепей: тяжелые Н (от англ. heavy - тяжелый) - 550-660
- 5. Домены различаются по постоянству аминокислотного состава. С-домены - постоянная структура полипептидной цепи, V-домены - переменная структура.
- 6. Молекулярное строение антител Н-цепь Fab
- 7. Дополнительные полипептидные цепи: J-пептид (от англ. join — соединяю) - обеспечивает превращение IgM и IgA в
- 8. В зависимости от особенностей молекулярного строения тяжелой цепи различают 5 классов иммуноглобулинов: IgA, IgG, IgM, IgE
- 9. Иммуноглобулины класса G. Составляют 70—80 % всех сывороточных Ig (12 г/л). Период полураспада IgG - 21
- 10. Иммуноглобулины класса G. Легко проходит через плацентарный барьер (обеспечивает гуморальный иммунитет новорожденного в первые 3—4 месяца
- 11. Иммуноглобулины класса М. Составляют 5-10 % всех сывороточных Ig (около 1 г/л). Период полураспада IgM -
- 12. Иммуноглобулины класса М. Участвует в формировании сывороточного и секреторного гуморального иммунитета. Не проходит через плаценту. Функции:
- 13. Иммуноглобулины класса А. Около 60 % всех IgA содержится в секретах слизистых. Сывороточный IgA Составляет 10—15
- 14. Секреторный IgA ди- или тример (4- или 6-валентный) содержит J- и S-пептиды. Синтезируется В-лимфоцитами и плазматическими
- 15. Иммуноглобулины класса Е. Содержание в сыворотке крови - около 0,002 %( 0,00025 г/л). Мономер, имеет 2
- 16. Иммуноглобулины класса D. Составляют 0,2 % от всех сывороточных Ig (около 0,03г/л). Мономер, имеет 2 антигенсвязывающих
- 17. Рецепторные (мембранные) иммуноглобулины. Локализуются на цитоплазматической мембране В-лимфоцитов. Выполняют функции антигенспецифических рецепторов. Структурное отличие - дополнительный
- 18. Нормальные (естественные) антитела. К ним относятся антитела, направленные против эритроцитарных антигенов групп крови (система АВО), антитела,
- 19. Моноклональные антитела - антитела с паратопом строго определенной специфичности. Такие антитела вырабатывает гибридома. Гибридома – гибридная
- 20. Полные антитела образуют в реакции агглютинации или преципитации (in vitro) хорошо различимую глазом макромолекулярную структуру гигантского
- 21. Взаимодействие антитела с антигеном Антигенсвязывающий центр, или паратоп (локализован в Fab-фрагменте) взаимодействует с антигенной детерминантой (эпитопом).
- 22. Нейтрализация - связывание и блокирование паратопом иммуноглобулина активного центра биологически активной молекулы (токсина, рецептора, лекарственного препарата
- 23. Фазы развития специфического иммунного ответа. Латентная фаза (скрытая, или индуктивная). Начинается с перестройки иммунной системы: происходит
- 24. Первичный ответ - иммунная реакция на первое воздействие антигена. Вторичный ответ - ответ на повторное воздействие
- 25. Первичный и вторичный иммунный ответ Уровень антител дни месяцы дни месяцы годы
- 26. Первичный и вторичный иммунный ответ Уровень антител Время (сутки)
- 27. Иммунологическая память способность организма при повторной встрече с антигеном формировать вторичный иммунный ответ. Имеет высокую специфичность
- 28. Два наиболее вероятных механизма формирования иммунологической памяти . 1. Длительное сохранение антигена в организме, что поддерживает
- 29. Феномен иммунологической памяти используется при вакцинации. Создается напряженный иммунитет и поддерживается длительное время на защитном уровне
- 31. Скачать презентацию

Основные формы проявления иммунитета:
антителообразование,
иммунный фагоцитоз,
опосредованный клетками киллинг,
реакции гиперчувствительности,
формирование
антителообразование,
иммунный фагоцитоз,
опосредованный клетками киллинг,
реакции гиперчувствительности,
формирование

Антитела (иммуноглобулины)
белки, специфически реагирующие с антигенами.
Относятся к гамма-глобулиновой фракции
белки, специфически реагирующие с антигенами.
Относятся к гамма-глобулиновой фракции

Молекулярное строение антител
2 пары полипептидных цепей:
тяжелые Н (от англ. heavy
Молекулярное строение антител
2 пары полипептидных цепей:
тяжелые Н (от англ. heavy

Домены различаются по постоянству аминокислотного состава.
С-домены - постоянная структура
С-домены - постоянная структура
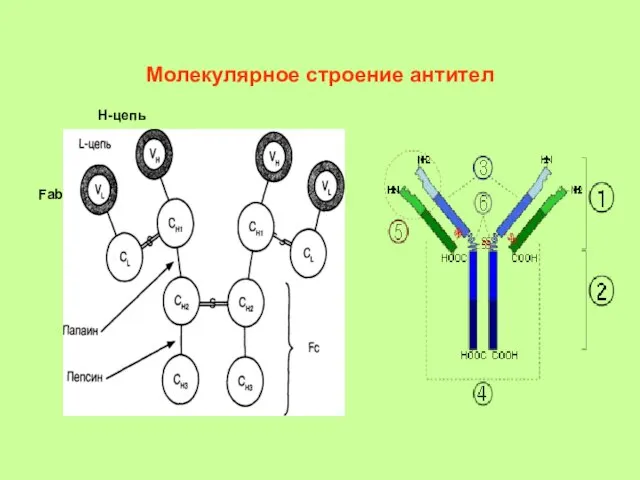
Молекулярное строение антител
Н-цепь
Fab
Молекулярное строение антител
Н-цепь
Fab
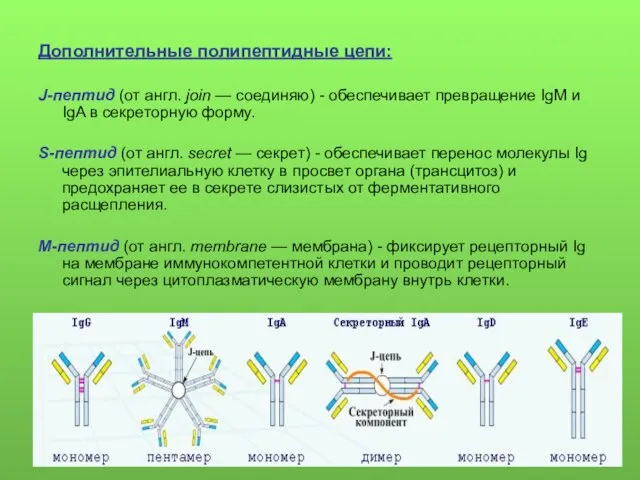
Дополнительные полипептидные цепи:
J-пептид (от англ. join — соединяю) - обеспечивает
Дополнительные полипептидные цепи:
J-пептид (от англ. join — соединяю) - обеспечивает
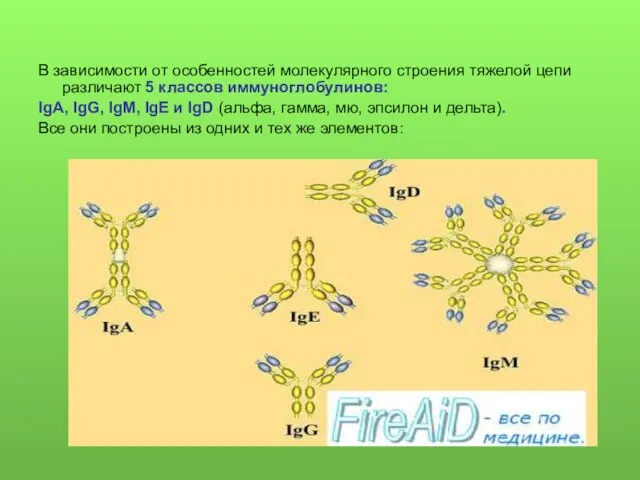
В зависимости от особенностей молекулярного строения тяжелой цепи различают 5 классов

Иммуноглобулины класса G.
Составляют 70—80 % всех сывороточных Ig (12 г/л).
Иммуноглобулины класса G.
Составляют 70—80 % всех сывороточных Ig (12 г/л).

Иммуноглобулины класса G.
Легко проходит через плацентарный барьер (обеспечивает гуморальный иммунитет
Иммуноглобулины класса G.
Легко проходит через плацентарный барьер (обеспечивает гуморальный иммунитет

Иммуноглобулины класса М.
Составляют 5-10 % всех сывороточных Ig (около 1
Иммуноглобулины класса М.
Составляют 5-10 % всех сывороточных Ig (около 1

Иммуноглобулины класса М.
Участвует в формировании сывороточного и секреторного гуморального иммунитета.
Иммуноглобулины класса М.
Участвует в формировании сывороточного и секреторного гуморального иммунитета.

Иммуноглобулины класса А.
Около 60 % всех IgA содержится в секретах
Иммуноглобулины класса А.
Около 60 % всех IgA содержится в секретах

Секреторный IgA
ди- или тример (4- или 6-валентный)
содержит J- и S-пептиды.
Секреторный IgA
ди- или тример (4- или 6-валентный)
содержит J- и S-пептиды.

Иммуноглобулины класса Е.
Содержание в сыворотке крови - около 0,002 %(
Содержание в сыворотке крови - около 0,002 %(

Иммуноглобулины класса D.
Составляют 0,2 % от всех сывороточных Ig (около
Составляют 0,2 % от всех сывороточных Ig (около

Рецепторные (мембранные) иммуноглобулины.
Локализуются на цитоплазматической мембране В-лимфоцитов.
Выполняют функции антигенспецифических
Рецепторные (мембранные) иммуноглобулины.
Локализуются на цитоплазматической мембране В-лимфоцитов.
Выполняют функции антигенспецифических

Нормальные (естественные) антитела.
К ним относятся
антитела, направленные против эритроцитарных антигенов групп крови
К ним относятся
антитела, направленные против эритроцитарных антигенов групп крови

Моноклональные антитела - антитела с паратопом строго определенной специфичности.
Такие антитела
Такие антитела

Полные антитела образуют в реакции агглютинации или преципитации (in vitro) хорошо
Полные антитела образуют в реакции агглютинации или преципитации (in vitro) хорошо

Взаимодействие антитела с антигеном
Антигенсвязывающий центр, или паратоп (локализован в Fab-фрагменте)
Взаимодействие антитела с антигеном
Антигенсвязывающий центр, или паратоп (локализован в Fab-фрагменте)

Нейтрализация - связывание и блокирование паратопом иммуноглобулина активного центра биологически активной

Фазы развития специфического иммунного ответа.
Латентная фаза (скрытая, или индуктивная). Начинается
Латентная фаза (скрытая, или индуктивная). Начинается

Первичный ответ - иммунная реакция на первое воздействие антигена.
Вторичный ответ
Вторичный ответ
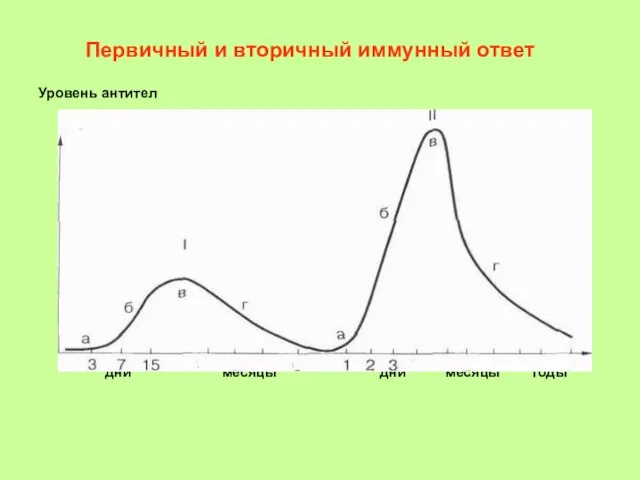
Первичный и вторичный иммунный ответ
Уровень антител
дни месяцы дни месяцы годы
Первичный и вторичный иммунный ответ
Уровень антител
дни месяцы дни месяцы годы
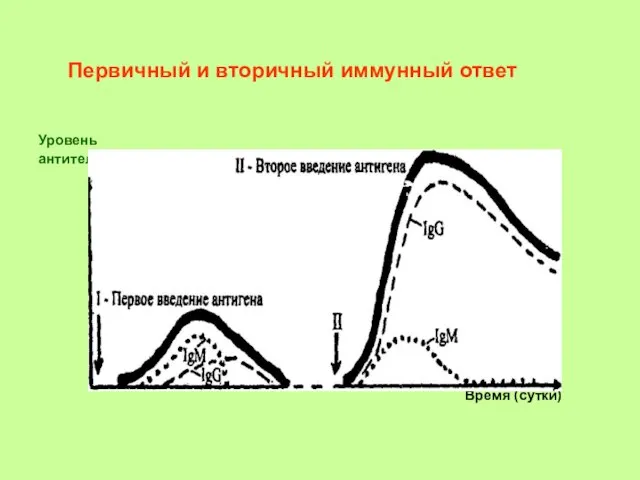
Первичный и вторичный иммунный ответ
Уровень
антител
Время (сутки)
Уровень
антител
Время (сутки)

Иммунологическая память
способность организма при повторной встрече с антигеном формировать вторичный
Иммунологическая память
способность организма при повторной встрече с антигеном формировать вторичный

Два наиболее вероятных механизма формирования иммунологической памяти .
1. Длительное
Два наиболее вероятных механизма формирования иммунологической памяти .
1. Длительное

Феномен иммунологической памяти
используется при вакцинации.
Создается напряженный иммунитет и
Феномен иммунологической памяти
используется при вакцинации.
Создается напряженный иммунитет и
 Инсульт. Түрлері мейірбикелік күтім
Инсульт. Түрлері мейірбикелік күтім Влияние характерологических особенностей на профессиональное самоопределение
Влияние характерологических особенностей на профессиональное самоопределение Кровотечения
Кровотечения Алынбайтын протездер
Алынбайтын протездер Медицинская помощь больным с менингитами на госпитальном этапе
Медицинская помощь больным с менингитами на госпитальном этапе Короткий световой день и организм человека
Короткий световой день и организм человека Финансирование здравоохранения в России
Финансирование здравоохранения в России Теоретические основы биомедицинской этики. Основные типы этических теорий
Теоретические основы биомедицинской этики. Основные типы этических теорий Бактериальная инфекция при инфекционном мононуклеозе
Бактериальная инфекция при инфекционном мононуклеозе Диссеменированые поражения легких. Редкие заболевания легких
Диссеменированые поражения легких. Редкие заболевания легких Психология личности
Психология личности Аутоиммунные болезни животных
Аутоиммунные болезни животных Противомикробные препараты требуют осторожного обращения
Противомикробные препараты требуют осторожного обращения О профилактике инфекций, передаваемых половым путем, или венерические болезни
О профилактике инфекций, передаваемых половым путем, или венерические болезни Синдром боли в животе
Синдром боли в животе Қант диабеті бар науқастарда ЗШЖИ
Қант диабеті бар науқастарда ЗШЖИ Синдром Клайнфельтера
Синдром Клайнфельтера Основы современной иммуногематологии
Основы современной иммуногематологии Синдром кошачьего крика
Синдром кошачьего крика Стентирование пищевода. Показания и противопоказания. Методика стентирования
Стентирование пищевода. Показания и противопоказания. Методика стентирования Введение в медицинскую психологию. Основные понятия. Классификации
Введение в медицинскую психологию. Основные понятия. Классификации Нарушения мышления
Нарушения мышления Иерсиниозы. Клиника и современные методы диагностики
Иерсиниозы. Клиника и современные методы диагностики Повреждения грудной клетки, живота, позвоночника и таза
Повреждения грудной клетки, живота, позвоночника и таза Эпидемический сыпной тиф. Болезнь Брилля-Цинссера
Эпидемический сыпной тиф. Болезнь Брилля-Цинссера Тұрғындардың ұжымдық және жекелей тамақтануының құндылығы
Тұрғындардың ұжымдық және жекелей тамақтануының құндылығы Нейродиетология (актуальное направление в детской неврологии)
Нейродиетология (актуальное направление в детской неврологии) Этика и деонтология фармацевтического работника
Этика и деонтология фармацевтического работника