- Эозинофилы. Базофилы. Тучные клетки

Содержание
- 2. Эозинофилы Подвижные, высоко дифференцированные клетки, 12—15 мкм в диаметре, имеют характерное двудольчатое ядро, развитый аппарат Гольджи
- 4. Мембранный белок
- 5. является молекулой адгезии, связывается с сиаловой кислотой Это высокоаффинный рецептор для следующих хемокинов: эотаксин (CCL11), эотаксин-3
- 6. Маркёр активации клеток гемопоэтической линии развития. Участвует во многих клеточных процессах, включая дифференцировку, адгезию и передачу
- 7. Созревание эозинофилов Эозинофилы происходят из СD34+ предшественников костного мозга. Созревание происходит под влиянием аутокринных и паракринных
- 8. Миграция эозинофилов В сосудистом русле эозинофилы пребывают в состоянии роллинга. Качение циркулирующих клеток по поверхности эндотелиоцитов
- 9. Хемотаксины эозинофилов
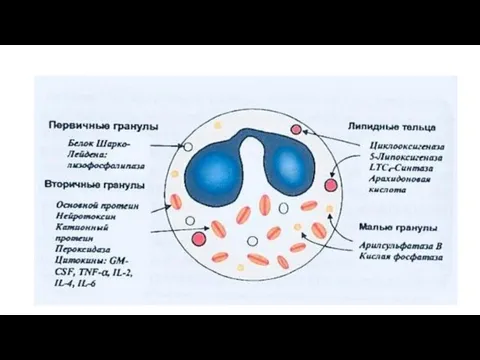
- 13. Гранулы эозинофилов
- 15. Секреция эозинофилов Выброс содержимого гранул за пределы клетки является основным проявлением реактивности эозинофила. Стадии: Мерокриновая секреция.
- 16. Частичная дегрануляция Этот процесс приводит к тому, что специфические гранулы начинают существенно различаться по морфологии: одни
- 17. Цитолиз Эозинофилы подвергаются цитолитической дегрануляции. При этом наблюдатся разрывы наружной и ядерной мембраны, лизис хроматина, набухание
- 18. Роль эозинофилов Осуществляют внеклеточный цитолиз, которому принадлежит основная роль в защите от многоклеточных паразитов. Большинство белков
- 19. Роль эозинофилов Свойственна слабая фагоцитарная активность. При активации в них образуются и затем секретируются разнообразные бактерицидные
- 20. Во время менструальных циклов и при беременности усиливается миграция эозинофилов в матку и молочные железы, где
- 21. Заболевания, связанные с эозинофилами
- 22. Базофилы Тканевая клетка, содержащая в цитоплазме базофильные гранулы. Диаметр 8-10мкм. При окрашивании по Романовскому- Гимза гранулы
- 23. CCR1 Хемокиновые рецепторы CCR2 CCR3 CXCR1 CXCR3 CXCR4 Взаимодействует с эотаксином (CCL11), эотаксин-3 (CCL26), MCP-3 (CCL7),
- 24. Онтогенез базофилов Предшественники базофилов с фенотипом Lin- CD34+FceRIhic-kit происходят из гранулоцитарно-макрофагальных предшественников костного мозга (GM-P) и
- 26. Активация базофилов IgE- зависимый путь активации базофилов. Базофилы экспрессируют высокоаффинный рецептор FceRI, который без участия АГ
- 27. IgE- независимый путь активации базофила. Существует несколько вариантов этого пути. Первый этап заключается в индукции образования
- 28. Первичные гранулы базофилов Относительно крупны . Мембрана гранул обладает высокой активностью фосфолипаз и липооксигеназы. Содержат: *пероксидазу
- 29. Вторичные мелкие гранулы Окружены более тонкой мембраной. В плазматической мембране базофилов имеются конституциональные высокоаффинные реагиновые рецепторы
- 30. Гранулы
- 32. Межклеточное взаимодействие 1. На ранних этапах развития иммунного ответа базофилы являются главными индукторами образования ТҺ2-клеток. 2.
- 33. Развитие и миграция тучных клеток и базофилов. Рядом с кружками, обозначающими клетки, указаны их маркеры. ОМЛ
- 34. Тучная клетка Диаметр тучных клеток варьирует от 10 до 20 мкм. Они имеют овальную форму с
- 35. Разновидности тучных клеток человека
- 37. Тучные клетки (1)— большие гранулярные клетки, происходят из CD34+ гемопоэтических предшественников костного мозга. Не циркулируют, а
- 38. FceR тучных клеток FceRI состоит из а-, в- и двух у-цепей, связанных между собой, а-цепь взаимодействует
- 39. Биологически активные вещества тучных клеток IgE (1), синтезируемый плазматическими клетками (2), при взаимодействии с рецептором FceRI
- 41. Функции тучных клеток
- 42. Функции тучных клеток Вещества, выделяемые ТК, вызывают развитие острого или хронического воспаления, лежащего в основе всех
- 46. Дегрануляция тучных клеток Две рядом лежащие молекулы IgE на поверхности мастоцита связываются 2 эпитопами молекулы поливалентного
- 51. Скачать презентацию
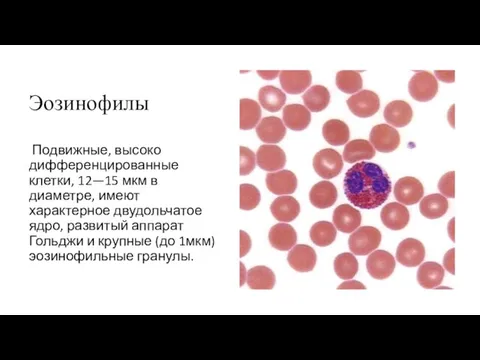
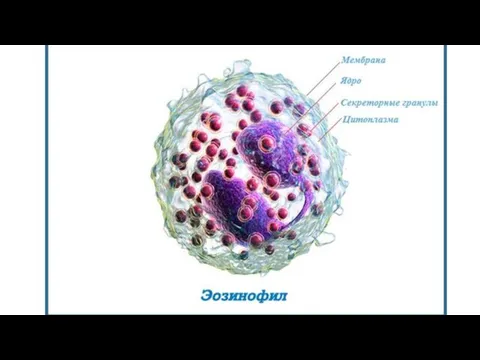
Эозинофилы
Подвижные, высоко дифференцированные клетки, 12—15 мкм в диаметре, имеют характерное
Эозинофилы
Подвижные, высоко дифференцированные клетки, 12—15 мкм в диаметре, имеют характерное
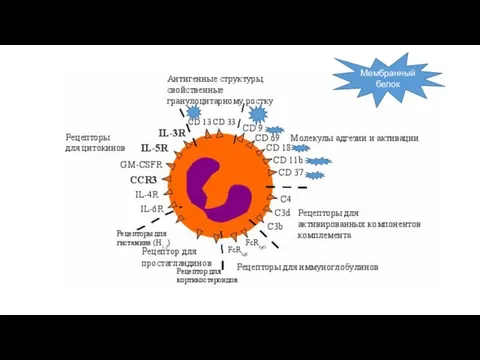
Мембранный белок
Мембранный белок

является молекулой адгезии, связывается с сиаловой кислотой
Это высокоаффинный рецептор для следующих
является молекулой адгезии, связывается с сиаловой кислотой
Это высокоаффинный рецептор для следующих
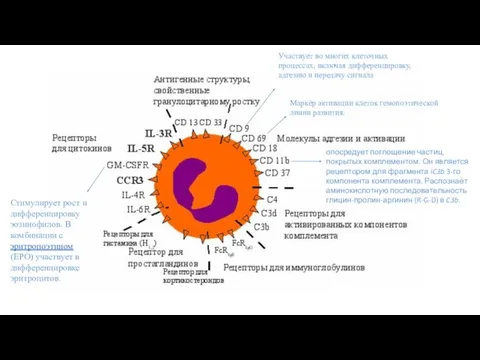
Маркёр активации клеток гемопоэтической линии развития.
Участвует во многих клеточных процессах, включая
Маркёр активации клеток гемопоэтической линии развития.
Участвует во многих клеточных процессах, включая
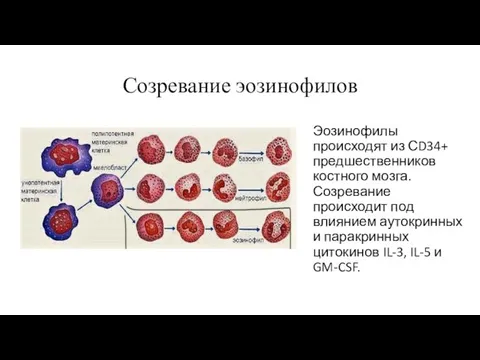
Созревание эозинофилов
Эозинофилы происходят из СD34+ предшественников костного мозга. Созревание происходит под
Созревание эозинофилов
Эозинофилы происходят из СD34+ предшественников костного мозга. Созревание происходит под

Миграция эозинофилов
В сосудистом русле эозинофилы пребывают в состоянии роллинга. Качение циркулирующих
Миграция эозинофилов
В сосудистом русле эозинофилы пребывают в состоянии роллинга. Качение циркулирующих
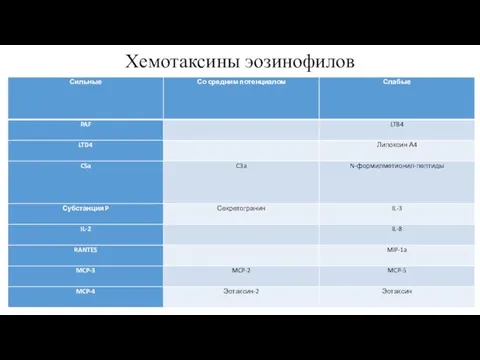
Хемотаксины эозинофилов
Хемотаксины эозинофилов



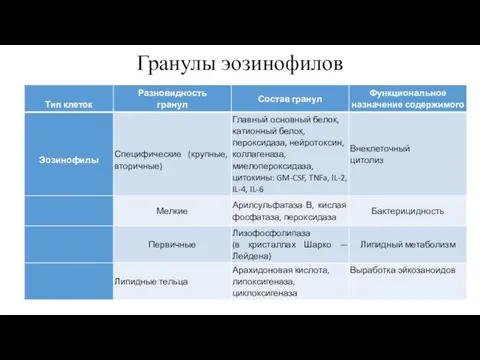
Гранулы эозинофилов
Гранулы эозинофилов

Секреция эозинофилов
Выброс содержимого гранул за пределы клетки является основным проявлением реактивности
Секреция эозинофилов
Выброс содержимого гранул за пределы клетки является основным проявлением реактивности

Частичная дегрануляция
Этот процесс приводит к тому, что специфические гранулы начинают
Частичная дегрануляция
Этот процесс приводит к тому, что специфические гранулы начинают

Цитолиз
Эозинофилы подвергаются цитолитической дегрануляции. При этом наблюдатся разрывы наружной и ядерной
Цитолиз
Эозинофилы подвергаются цитолитической дегрануляции. При этом наблюдатся разрывы наружной и ядерной

Роль эозинофилов
Осуществляют внеклеточный цитолиз, которому принадлежит основная роль в защите от
Роль эозинофилов
Осуществляют внеклеточный цитолиз, которому принадлежит основная роль в защите от

Роль эозинофилов
Свойственна слабая фагоцитарная активность. При активации в них образуются и
Роль эозинофилов
Свойственна слабая фагоцитарная активность. При активации в них образуются и

Во время менструальных циклов и при беременности усиливается миграция эозинофилов в
Во время менструальных циклов и при беременности усиливается миграция эозинофилов в
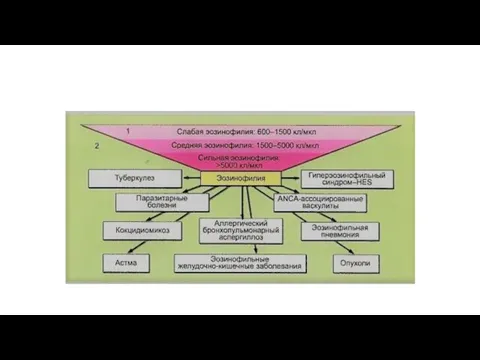
Заболевания, связанные с эозинофилами
Заболевания, связанные с эозинофилами
Базофилы
Тканевая клетка, содержащая в цитоплазме базофильные гранулы. Диаметр 8-10мкм. При окрашивании
Базофилы
Тканевая клетка, содержащая в цитоплазме базофильные гранулы. Диаметр 8-10мкм. При окрашивании
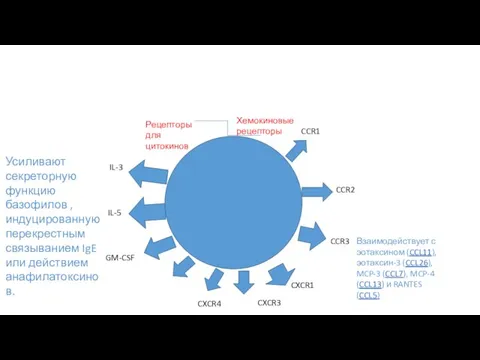
CCR1
Хемокиновые рецепторы
CCR2
CCR3
CXCR1
CXCR3
CXCR4
Взаимодействует с эотаксином (CCL11), эотаксин-3 (CCL26), MCP-3 (CCL7), MCP-4 (CCL13)
CCR1
Хемокиновые рецепторы
CCR2
CCR3
CXCR1
CXCR3
CXCR4
Взаимодействует с эотаксином (CCL11), эотаксин-3 (CCL26), MCP-3 (CCL7), MCP-4 (CCL13)
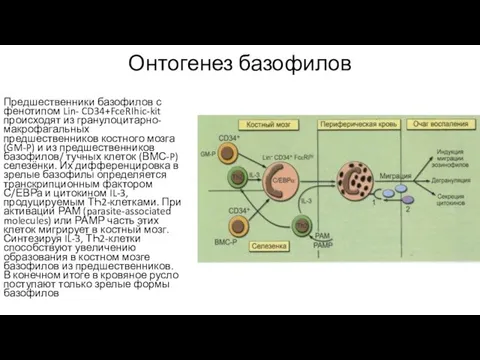
Онтогенез базофилов
Предшественники базофилов с фенотипом Lin- CD34+FceRIhic-kit происходят из гранулоцитарно-макрофагальных предшественников
Онтогенез базофилов
Предшественники базофилов с фенотипом Lin- CD34+FceRIhic-kit происходят из гранулоцитарно-макрофагальных предшественников
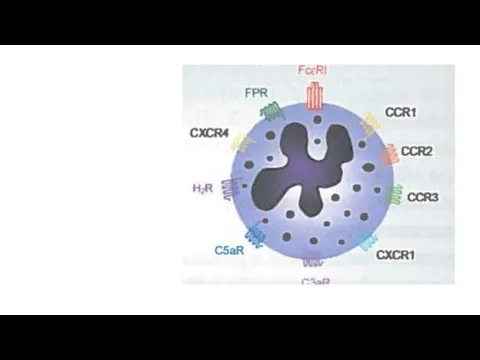

Активация базофилов
IgE- зависимый путь активации базофилов. Базофилы экспрессируют высокоаффинный рецептор
Активация базофилов
IgE- зависимый путь активации базофилов. Базофилы экспрессируют высокоаффинный рецептор

IgE- независимый путь активации базофила. Существует несколько вариантов этого пути. Первый
IgE- независимый путь активации базофила. Существует несколько вариантов этого пути. Первый

Первичные гранулы базофилов
Относительно крупны . Мембрана гранул обладает высокой активностью фосфолипаз
Первичные гранулы базофилов
Относительно крупны . Мембрана гранул обладает высокой активностью фосфолипаз

Вторичные мелкие гранулы
Окружены более тонкой мембраной.
В плазматической мембране базофилов имеются
Вторичные мелкие гранулы
Окружены более тонкой мембраной.
В плазматической мембране базофилов имеются
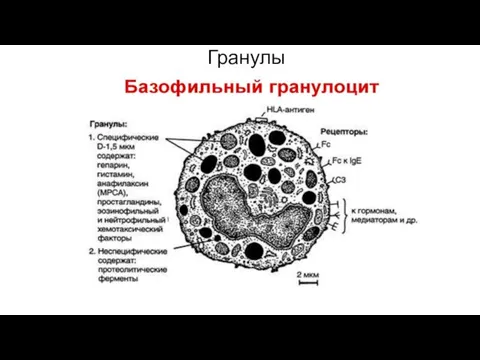
Гранулы
Гранулы


Межклеточное взаимодействие
1. На ранних этапах развития иммунного ответа базофилы являются главными
Межклеточное взаимодействие
1. На ранних этапах развития иммунного ответа базофилы являются главными

Развитие и миграция тучных клеток и базофилов.
Рядом с кружками, обозначающими
Развитие и миграция тучных клеток и базофилов.
Рядом с кружками, обозначающими
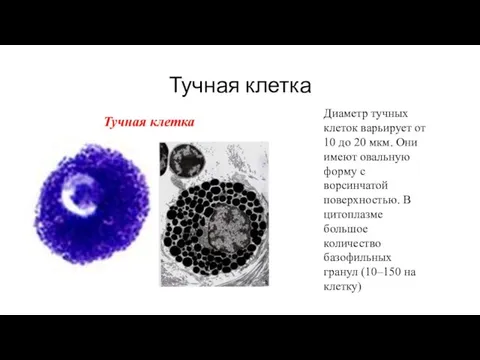
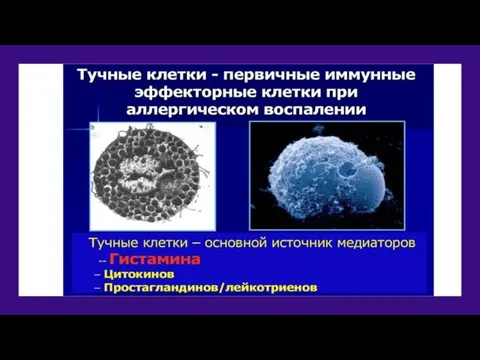
Тучная клетка
Диаметр тучных клеток варьирует от 10 до 20 мкм. Они
Тучная клетка
Диаметр тучных клеток варьирует от 10 до 20 мкм. Они

Разновидности тучных клеток человека
Разновидности тучных клеток человека

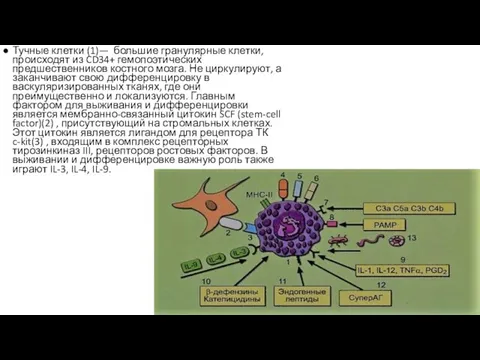
Тучные клетки (1)— большие гранулярные клетки, происходят из CD34+ гемопоэтических предшественников
Тучные клетки (1)— большие гранулярные клетки, происходят из CD34+ гемопоэтических предшественников

FceR тучных клеток
FceRI состоит из а-, в- и двух у-цепей, связанных
FceR тучных клеток
FceRI состоит из а-, в- и двух у-цепей, связанных

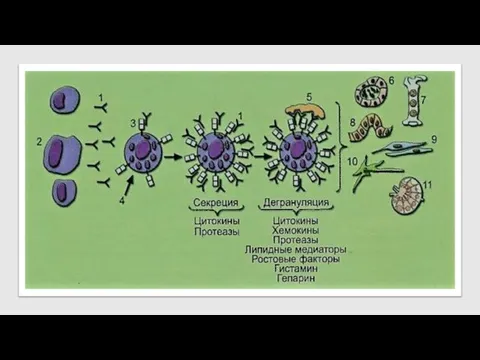
Биологически активные вещества тучных клеток
IgE (1), синтезируемый плазматическими клетками (2), при
Биологически активные вещества тучных клеток
IgE (1), синтезируемый плазматическими клетками (2), при
Функции тучных клеток
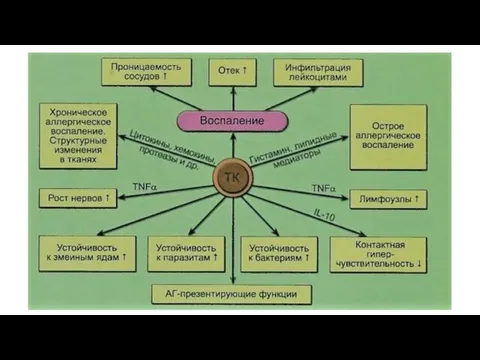
Функции тучных клеток

Функции тучных клеток
Вещества, выделяемые ТК, вызывают развитие острого или хронического
Функции тучных клеток
Вещества, выделяемые ТК, вызывают развитие острого или хронического



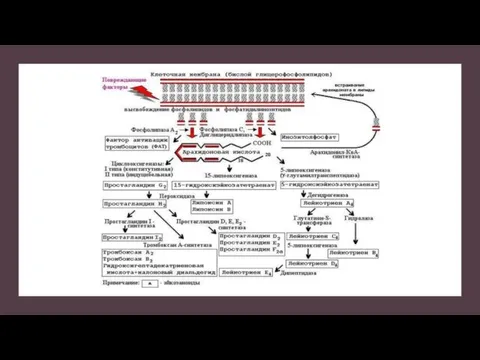
Дегрануляция тучных клеток
Две рядом лежащие молекулы IgE на поверхности мастоцита связываются
Дегрануляция тучных клеток
Две рядом лежащие молекулы IgE на поверхности мастоцита связываются

 Дермоид роговицы или дермоидная киста у собак и кошек
Дермоид роговицы или дермоидная киста у собак и кошек Черепно-мозговая травма
Черепно-мозговая травма Отхаркивающие средства
Отхаркивающие средства Показания к ЭФИ. Электрофизиологическое лечение ХСН
Показания к ЭФИ. Электрофизиологическое лечение ХСН Синдром Дауна
Синдром Дауна Крапивинская районная больница. Приглашаем на работу
Крапивинская районная больница. Приглашаем на работу Влияние электромагнитных волн на организм
Влияние электромагнитных волн на организм Общая характеристика лекарственных растений
Общая характеристика лекарственных растений Протезирование органов чувств (лекция 2)
Протезирование органов чувств (лекция 2) Сестринская помощь и эпидемический процесс
Сестринская помощь и эпидемический процесс Болезни кожи с невыясненной этиологией
Болезни кожи с невыясненной этиологией Воспалительные заболевания органов репродуктивной системы женщины
Воспалительные заболевания органов репродуктивной системы женщины Хирургическая операция
Хирургическая операция Гемолитическая желтуха новорожденных
Гемолитическая желтуха новорожденных Жеке адамның индивидуалды ерекшеліктерін талдау және оны тергеу, сот, сауықтыру және түзету жұмыстарында есепке алу
Жеке адамның индивидуалды ерекшеліктерін талдау және оны тергеу, сот, сауықтыру және түзету жұмыстарында есепке алу Горная болезнь
Горная болезнь Қазақстандағы денсаулық сақтау ұйымы және бағдарламалары
Қазақстандағы денсаулық сақтау ұйымы және бағдарламалары Сахар, аминокислоты, соли, минералы
Сахар, аминокислоты, соли, минералы Урок математики 4 класс учитель начальных классов МБОУ СОШ№ 156 г. Новосибирск Черемисина С.В.
Урок математики 4 класс учитель начальных классов МБОУ СОШ№ 156 г. Новосибирск Черемисина С.В. Правильное питание – залог здоровья
Правильное питание – залог здоровья Плазменные факторы свертывания крови (плазменные прокоагулянты). Геморрагические диатезы
Плазменные факторы свертывания крови (плазменные прокоагулянты). Геморрагические диатезы Рисуночный тест Гилфорда «Социальный интеллект»
Рисуночный тест Гилфорда «Социальный интеллект» Русская баня и ее полезные свойства
Русская баня и ее полезные свойства Modulis skolēnu izziņas aktivitātes pedagoģiskie un psiholoģiskie pamati
Modulis skolēnu izziņas aktivitātes pedagoģiskie un psiholoģiskie pamati Сухожильный шов
Сухожильный шов Варианты ответа миокарда на ишемию
Варианты ответа миокарда на ишемию Биосоциологическая теория (Ч. Ломброзо, У. Шелдон)
Биосоциологическая теория (Ч. Ломброзо, У. Шелдон) Рациональное использование лекарственных средств при неотложных состояниях
Рациональное использование лекарственных средств при неотложных состояниях