- Клеточный тип иммунного ответа

Содержание
- 2. Введение. Клеточный иммунитет (англ. Cell-mediated immunity) — такой тип иммунного ответа, в котором не участвуют ни
- 3. Клеточный иммунный ответ. Типы эффекторных Т-клеток и их функции. Взаимодействие адаптивных Т-клеток с эффекторами или мишенями,
- 4. Цитотоксический Т-клеточный иммунный ответ.
- 5. Цитотоксический Т-клеточный иммунный ответ. Цитотоксический иммунный ответ осуществляют Т-лимфоциты, экспрессирующие корецептор CD8. Это определяет главную особенность
- 6. Естественные и индуцированные цитотоксические Т-лимфоциты.
- 7. Цитотоксический иммунный ответ проходит в 4этапа. I. Презентация II. Пролиферация III. Дифференцировка IV. Цитолиз антигена клеток
- 8. Распознавание антигенного пептида и активация CD8 Т-клеток. Вирусом может быть инфицирована практически любая клетка организма. Однако
- 9. Распознавание антигенного пептида и активация CD8 Т-клеток. Особенности распознавания антигена при индукции развития цитотоксических Т-лимфоцитов и
- 10. Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток. Долгое время участие CD4+ Т-хелперов в развитии цитотоксического
- 11. Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток. Именно степень самообеспечения активированных CD8+ Т-клеток аутокринным ростовым
- 12. Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток. Таким образом, хотя CD8+ Т-клетки, вовлекаемые в цитотоксический
- 13. Цитотоксические Т-лимфоциты. Как и в случае Т-хелперов, дифференцировка цитотоксических Т-лимфоцитов начинается в процессе их пролиферативной экспансии.
- 14. Цитотоксические Т-лимфоциты. В процессе дифференцировки в эффекторные клетки происходит сплайсинг участков РНК, кодируемых экзонами сначала А,
- 15. Иммунный Т-клеточный цитолиз. Механизмы реализации цитотоксического эффекта через индукцию апоптоза. Цитолиз клеток-мишеней цитотоксическими Т-лимфоцитами осуществляется с
- 16. Иммунный Т-клеточный цитолиз. Как и при цитолизе, осуществляемом естественными киллерами, между цитотоксическим Т-лимфоцитом и клеткой-мишенью формируется
- 17. Иммунный Т-клеточный цитолиз. Перфорин, поступающий в микрополость, в присутствии ионов Ca2+ изменяет свою конформацию: на поверхности
- 18. Fas-зависимый цитолиз. Цитотоксические Т-лимфоциты используют еще один механизм контактного киллинга, причем в большей степени, чем естественные
- 19. Миграция клеток при цитотоксическом иммунном ответе. Цитотоксический клеточный иммунный ответ участвует преимущественно в защите от вирусных
- 20. Миграция клеток при цитотоксическом иммунном ответе. В очагах инфицирования вирусами и другими патогенами цитотоксические Т-лимфоциты реализуют
- 21. Миграция клеток при цитотоксическом иммунном ответе. После успешного завершения цитотоксического иммунного ответа происходит быстрая и радикальная
- 22. Воспалительный Т-клеточный иммунный ответ.
- 23. Воспалительный Т-клеточный иммунный ответ. Эта форма иммунного ответа предназначена для защиты от внутриклеточных патогенов, локализующихся в
- 24. Воспалительный Т-клеточный иммунный ответ. Схема развития клеточного иммунного ответа воспалительного типа. Клеточный иммунный ответ воспалительного типа
- 25. Воспалительный Т-клеточный иммунный ответ. За реализацию этой формы защиты отвечают Thl-клетки и макрофаги. Thl-клетки формируются на
- 26. Активирующее взаимодействие Thl-клеток с макрофагами. Этот этап характерен именно для воспалительного иммунного ответа. Он состоит во
- 27. Активирующее взаимодействие Thl-клеток с макрофагами. Активация макрофагов Т-хелперами. Показаны механизмы воздействия Thl-клеток на макрофаги (контактный и
- 28. Активирующее взаимодействие Thl-клеток с макрофагами. Роль интерферона у в дифференцировке и реализации функций Т-хелперов. Интерферон, продуцируемый
- 29. Воспалительная составляющая Thl-клеточного иммунного ответа. В отличие от цитотоксического иммунного ответа, не связанного очевидным образом с
- 30. Иммунное воспаление. Вариант воспаления, реализуемый с участием Th1-клеток, называют иммунным воспалением, а сам Th1-клеточный иммунный ответ
- 32. Скачать презентацию

Введение.
Клеточный иммунитет (англ. Cell-mediated immunity) — такой тип иммунного ответа, в
Введение.
Клеточный иммунитет (англ. Cell-mediated immunity) — такой тип иммунного ответа, в

Клеточный иммунный ответ.
Типы эффекторных Т-клеток и их функции. Взаимодействие адаптивных Т-клеток
Клеточный иммунный ответ.
Типы эффекторных Т-клеток и их функции. Взаимодействие адаптивных Т-клеток

Цитотоксический Т-клеточный иммунный ответ.
Цитотоксический Т-клеточный иммунный ответ.

Цитотоксический Т-клеточный иммунный ответ.
Цитотоксический иммунный ответ осуществляют Т-лимфоциты, экспрессирующие корецептор CD8.
Цитотоксический Т-клеточный иммунный ответ.
Цитотоксический иммунный ответ осуществляют Т-лимфоциты, экспрессирующие корецептор CD8.
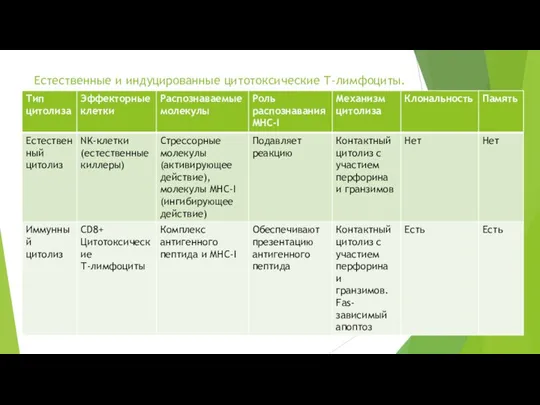
Естественные и индуцированные цитотоксические Т-лимфоциты.
Естественные и индуцированные цитотоксические Т-лимфоциты.

Цитотоксический иммунный ответ проходит в 4этапа.
I. Презентация II. Пролиферация III. Дифференцировка
Цитотоксический иммунный ответ проходит в 4этапа.
I. Презентация II. Пролиферация III. Дифференцировка

Распознавание антигенного пептида и активация CD8 Т-клеток.
Вирусом может быть инфицирована практически
Распознавание антигенного пептида и активация CD8 Т-клеток.
Вирусом может быть инфицирована практически

Распознавание антигенного пептида и активация CD8 Т-клеток.
Особенности распознавания антигена при индукции
Распознавание антигенного пептида и активация CD8 Т-клеток.
Особенности распознавания антигена при индукции

Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток.
Долгое время участие CD4+
Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток.
Долгое время участие CD4+

Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток.
Именно степень самообеспечения активированных
Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток.
Именно степень самообеспечения активированных

Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток.
Таким образом, хотя CD8+
Роль Т-хелперов и IL-2 в ответе CD8 Т-клеток.
Таким образом, хотя CD8+

Цитотоксические Т-лимфоциты.
Как и в случае Т-хелперов, дифференцировка цитотоксических Т-лимфоцитов начинается в
Цитотоксические Т-лимфоциты.
Как и в случае Т-хелперов, дифференцировка цитотоксических Т-лимфоцитов начинается в

Цитотоксические Т-лимфоциты.
В процессе дифференцировки в эффекторные клетки происходит сплайсинг участков РНК,
Цитотоксические Т-лимфоциты.
В процессе дифференцировки в эффекторные клетки происходит сплайсинг участков РНК,

Иммунный Т-клеточный цитолиз.
Механизмы реализации цитотоксического эффекта через индукцию апоптоза.
Цитолиз клеток-мишеней цитотоксическими
Иммунный Т-клеточный цитолиз.
Механизмы реализации цитотоксического эффекта через индукцию апоптоза.
Цитолиз клеток-мишеней цитотоксическими

Иммунный Т-клеточный цитолиз.
Как и при цитолизе, осуществляемом естественными киллерами, между цитотоксическим
Иммунный Т-клеточный цитолиз.
Как и при цитолизе, осуществляемом естественными киллерами, между цитотоксическим

Иммунный Т-клеточный цитолиз.
Перфорин, поступающий в микрополость, в присутствии ионов Ca2+ изменяет
Иммунный Т-клеточный цитолиз.
Перфорин, поступающий в микрополость, в присутствии ионов Ca2+ изменяет

Fas-зависимый цитолиз.
Цитотоксические Т-лимфоциты используют еще один механизм контактного киллинга, причем в
Fas-зависимый цитолиз.
Цитотоксические Т-лимфоциты используют еще один механизм контактного киллинга, причем в

Миграция клеток при цитотоксическом иммунном ответе.
Цитотоксический клеточный иммунный ответ участвует преимущественно
Миграция клеток при цитотоксическом иммунном ответе.
Цитотоксический клеточный иммунный ответ участвует преимущественно

Миграция клеток при цитотоксическом иммунном ответе.
В очагах инфицирования вирусами и другими
Миграция клеток при цитотоксическом иммунном ответе.
В очагах инфицирования вирусами и другими

Миграция клеток при цитотоксическом иммунном ответе.
После успешного завершения цитотоксического иммунного ответа
Миграция клеток при цитотоксическом иммунном ответе.
После успешного завершения цитотоксического иммунного ответа

Воспалительный Т-клеточный иммунный ответ.
Воспалительный Т-клеточный иммунный ответ.

Воспалительный Т-клеточный иммунный ответ.
Эта форма иммунного ответа предназначена для защиты от
Воспалительный Т-клеточный иммунный ответ.
Эта форма иммунного ответа предназначена для защиты от

Воспалительный Т-клеточный иммунный ответ.
Схема развития клеточного иммунного ответа воспалительного типа.
Клеточный иммунный
Воспалительный Т-клеточный иммунный ответ.
Схема развития клеточного иммунного ответа воспалительного типа.
Клеточный иммунный

Воспалительный Т-клеточный иммунный ответ.
За реализацию этой формы защиты отвечают Thl-клетки и
Воспалительный Т-клеточный иммунный ответ.
За реализацию этой формы защиты отвечают Thl-клетки и

Активирующее взаимодействие Thl-клеток с макрофагами.
Этот этап характерен именно для воспалительного иммунного
Активирующее взаимодействие Thl-клеток с макрофагами.
Этот этап характерен именно для воспалительного иммунного

Активирующее взаимодействие Thl-клеток с макрофагами.
Активация макрофагов Т-хелперами. Показаны механизмы воздействия Thl-клеток
Активирующее взаимодействие Thl-клеток с макрофагами.
Активация макрофагов Т-хелперами. Показаны механизмы воздействия Thl-клеток
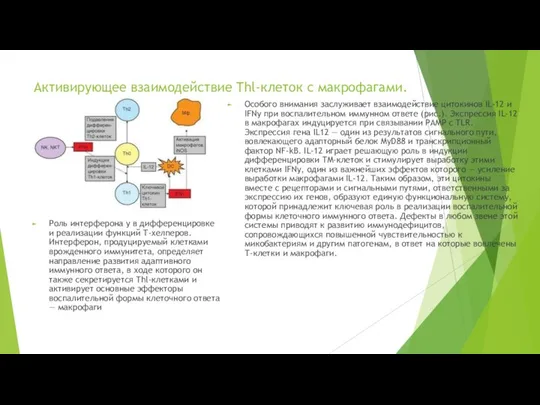
Активирующее взаимодействие Thl-клеток с макрофагами.
Роль интерферона у в дифференцировке и реализации
Активирующее взаимодействие Thl-клеток с макрофагами.
Роль интерферона у в дифференцировке и реализации

Воспалительная составляющая Thl-клеточного иммунного ответа.
В отличие от цитотоксического иммунного ответа, не
Воспалительная составляющая Thl-клеточного иммунного ответа.
В отличие от цитотоксического иммунного ответа, не

Иммунное воспаление.
Вариант воспаления, реализуемый с участием Th1-клеток, называют иммунным воспалением, а
Иммунное воспаление.
Вариант воспаления, реализуемый с участием Th1-клеток, называют иммунным воспалением, а
 Паратозилярлы абцесс
Паратозилярлы абцесс Особенности влияния различных заболеваний на эмоциональную сферу взрослых и детей
Особенности влияния различных заболеваний на эмоциональную сферу взрослых и детей Онкогенные вирусы. Особенности противоопухолевого иммунитета
Онкогенные вирусы. Особенности противоопухолевого иммунитета Коклюш у детей
Коклюш у детей Бронхоскопия. Показания. Противопоказания
Бронхоскопия. Показания. Противопоказания Физическая реабилитация при сколиозе II степени у детей 14-16 лет
Физическая реабилитация при сколиозе II степени у детей 14-16 лет Первая помощь при кровотечениях
Первая помощь при кровотечениях Питание ребенка дошкольного возраста (4-7 лет)
Питание ребенка дошкольного возраста (4-7 лет) Справка по заполнению формы «Перечень обособленных подразделений ЛПУ (без ФАП и ФП), требующих подключения к сети Интернет
Справка по заполнению формы «Перечень обособленных подразделений ЛПУ (без ФАП и ФП), требующих подключения к сети Интернет Факторы неспецифической и специфической защиты организма. Понятие об антигенах и антителах. Иммунокомпетентные клетки
Факторы неспецифической и специфической защиты организма. Понятие об антигенах и антителах. Иммунокомпетентные клетки Психологические и этические аспекты профессиональной деятельности фармацевтов
Психологические и этические аспекты профессиональной деятельности фармацевтов Функциональная анатомия височно-нижнечелюстного сустава (ВНЧС)
Функциональная анатомия височно-нижнечелюстного сустава (ВНЧС) Гемолитико-уремический синдром (болезнь Гассера)
Гемолитико-уремический синдром (болезнь Гассера) Комп'ютерна залежність - ознаки, стадії, причини виникнення та профілактика. Селфіманія - хвороба чи спосіб самовираження
Комп'ютерна залежність - ознаки, стадії, причини виникнення та профілактика. Селфіманія - хвороба чи спосіб самовираження Патология терморегуляции. Гипертермия и лихорадка. Стадии, виды, значение
Патология терморегуляции. Гипертермия и лихорадка. Стадии, виды, значение Что такое страх
Что такое страх Психология
Психология Психологія колективу
Психологія колективу Эргономические основы массажа
Эргономические основы массажа Постинсультная депрессия и деменция
Постинсультная депрессия и деменция Несбалансированное питание и развитие костно–суставных заболеваний
Несбалансированное питание и развитие костно–суставных заболеваний Язвенная болезнь желудка и двенадцатиперстной кишки
Язвенная болезнь желудка и двенадцатиперстной кишки Гигиеническая и хирургическая обработка рук современными методами
Гигиеническая и хирургическая обработка рук современными методами Управление конфликтами. Тема 10
Управление конфликтами. Тема 10 Введение в рентгенологию. Лекция №1
Введение в рентгенологию. Лекция №1 Аллергические реакции
Аллергические реакции Дисграфия
Дисграфия Психолого-педагогическая характеристика детей с особыми образовательными потребностями
Психолого-педагогическая характеристика детей с особыми образовательными потребностями