Скелетная мышца: строение мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы
- Скелетная мышца: строение мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы

Содержание
- 2. Ультраструктура скелетной мышцы. Поперечно-полосатая мышца состоит из множества функциональных единиц – мышечных волокон или мышечных клеток.
- 3. Поперек волокна между миофибриллами проходит система трубочек, представляющих впячивания сарколеммы, называемые Т-системой. В определенных местах трубочки
- 4. Молекула миозина состоит из длинного участка – хвоста и глобулярной части из двух головок. Молекулы миозина
- 5. Рис. 6. Схематическое изображение тонкой актиновой нити , показывающее локализацию тропомиозина и тропонина на его поверхности.
- 6. Тропонин участвует в Са2+ - зависимой регуляции сокращения. Тропонин Т имеет участок для связывания тропомиозина и
- 7. Актиновые филаменты имеют сложный состав и структуру. Каждая актиновая нить состоит из двух спиральных тяжей, каждый
- 8. Типы мышечных волокон. По способу ответа на возбуждающие нервные импульсы скелетные поперечно-полосатые мышцы подразделяют на тонические
- 9. Фазные волокна, наоборот, сокращаются по принципу «все или ничего». Как только нервный импульс достигнет надпороговой силы,
- 10. Фазные волокна по скорости развития напряжения подразделяют на «медленные» – волокна I типа и «быстрые» –
- 11. Проф. Коц Я.М.
- 12. 4-2. СОВРЕМЕННАЯ ТЕОРИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ. ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ. Механизм мышечного сокращения. В присутствии АТФ, но
- 13. Цикл после завершения может повториться или остановиться. Если бы все поперечные мостики в отдельной мышце прошли
- 14. Электромеханическое сопряжение. Трансформация возбуждения в сокращение, по существу представляющая передачу сигнала о начале сокращения от возбужденной
- 15. Остановка сокращения Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного импульса в двигательной концевой пластинке
- 16. 4-3. ВИДЫ И РЕЖИМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ. ОДИНОЧНОЕ СОКРАЩЕНИЕ. ПОНЯТИЕ О ТЕТАНУСЕ. МЕХАНИЗМ ДЛИТЕЛЬНОГО СОКРАЩЕНИЯ В ЕСТЕСТВЕННЫХ
- 17. Когда мышца укорачивается, преодолевая постоянную нагрузку, говорят об изотоническом сокращении. Большинство сокращений в организме представляют комбинацию
- 18. Одиночное сокращение и тетанус. Одиночный стимул спустя короткое время, около 50 мс, называемое латентным периодом, вызывает
- 19. Если следующий нервный импульс приходит к волокну раньше, чем закончилось расслабление предыдущего одиночного сокращения, происходит суперпозиция
- 20. При достаточно высокой частоте стимуляции выходящий из ретикулума Са2+ не успевает вернуться в цистерны, а мышца
- 21. Сила сокращения скелетных мышц. Двигательная единица. Под двигательной единицей понимают каждую группу мышечных волокон вместе с
- 24. Максимальная сила мышц. Абсолютная сила, развиваемая разными мышцами, неодинакова. Она определяется: 1) поперечным сечением мышцы, 2)
- 26. 4.4. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И СВОЙСТВА гладких мышц. Классификация гладких мышц. Висцеральные гладкие мышцы находятся во всех
- 28. Иннервация гладких мышц. Висцеральная гладкая мышца имеет двойную иннервацию — симпатическую и парасимпатическую, функция которой заключается
- 30. Нервно-мышечная передача. Возбуждающее влияние адренергических или холинергических нервов электрически проявляется в виде отдельных волн деполяризации. При
- 31. В периоды состояния относительного покоя величина мембранного потенциала в среднем равна — 50 мВ. В клетках
- 32. Автоматия. Многим гладким мышцам свойственна спонтанная, автоматическая активность. Для нее характерно медленное снижение мембранного потенциала покоя,
- 33. Механизм сокращения. В механизме сокращения гладкой мышцы имеется особенность, отличающая его от механизма сокращения скелетной мышцы.
- 36. Химическая чувствительность. Гладкие мышцы обладают высокой чувствительностью к различным физиологически активным веществам: адреналину, норадреналину, АХ, гистамину
- 37. Проведение возбуждения по гладкой мышце. В отличие от того, что имеет место в скелетных мышцах, в
- 38. Так же, как и в скелетной мускулатуре, в гладкой потенциалы действия имеют пусковое значение для начала
- 39. Вследствие медленности сокращения гладкая мышца даже при редких ритмических раздражениях (10-12 в мин) легко переходит в
- 41. Скачать презентацию
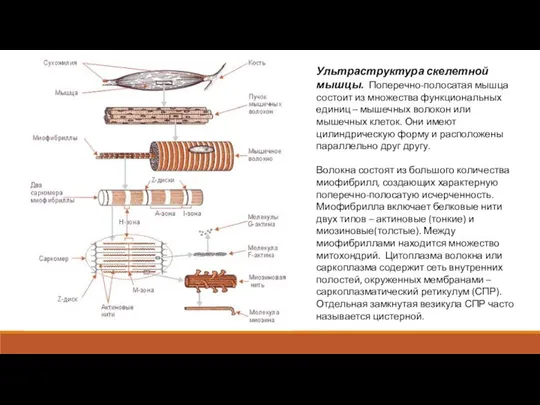
Ультраструктура скелетной мышцы. Поперечно-полосатая мышца состоит из множества функциональных единиц –
Ультраструктура скелетной мышцы. Поперечно-полосатая мышца состоит из множества функциональных единиц –

Поперек волокна между миофибриллами проходит система трубочек, представляющих впячивания сарколеммы, называемые
Поперек волокна между миофибриллами проходит система трубочек, представляющих впячивания сарколеммы, называемые
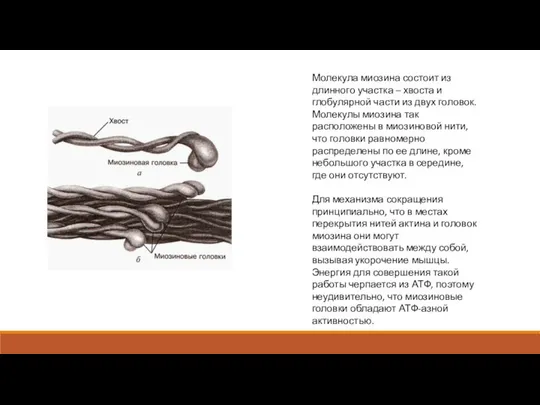
Молекула миозина состоит из длинного участка – хвоста и глобулярной части
Молекула миозина состоит из длинного участка – хвоста и глобулярной части
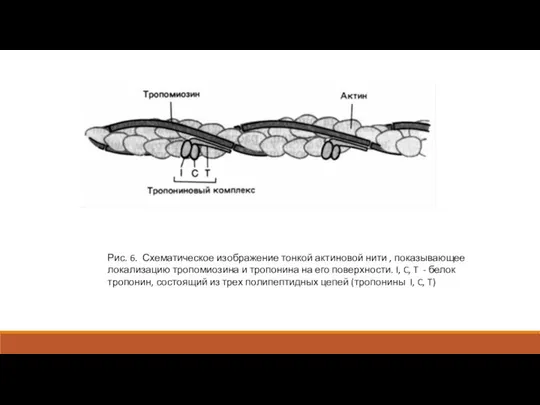
Рис. 6. Схематическое изображение тонкой актиновой нити , показывающее локализацию тропомиозина
Рис. 6. Схематическое изображение тонкой актиновой нити , показывающее локализацию тропомиозина
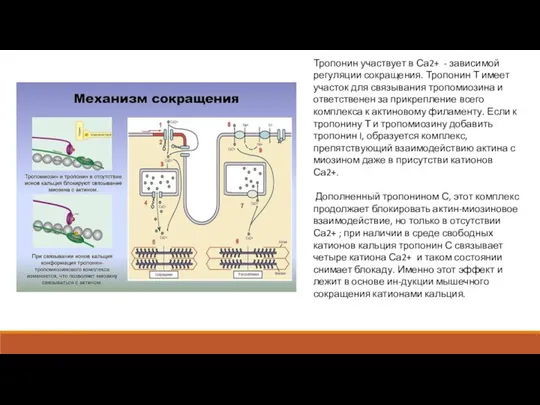
Тропонин участвует в Са2+ - зависимой регуляции сокращения. Тропонин Т имеет
Тропонин участвует в Са2+ - зависимой регуляции сокращения. Тропонин Т имеет
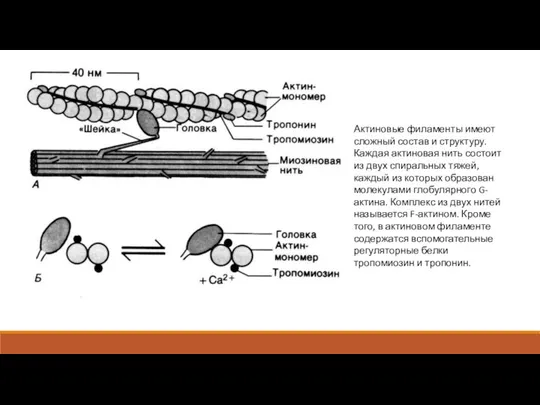
Актиновые филаменты имеют сложный состав и структуру. Каждая актиновая нить состоит
Актиновые филаменты имеют сложный состав и структуру. Каждая актиновая нить состоит

Типы мышечных волокон. По способу ответа на возбуждающие нервные импульсы скелетные
Типы мышечных волокон. По способу ответа на возбуждающие нервные импульсы скелетные

Фазные волокна, наоборот, сокращаются по принципу «все или ничего». Как только
Фазные волокна, наоборот, сокращаются по принципу «все или ничего». Как только

Фазные волокна по скорости развития напряжения подразделяют на «медленные» – волокна
Фазные волокна по скорости развития напряжения подразделяют на «медленные» – волокна

Проф.
Коц Я.М.
Проф.
Коц Я.М.

4-2. СОВРЕМЕННАЯ ТЕОРИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ.
ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ.
Механизм мышечного
4-2. СОВРЕМЕННАЯ ТЕОРИЯ МЫШЕЧНОГО СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ.
ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ.
Механизм мышечного

Цикл после завершения может повториться или остановиться. Если бы все поперечные
Цикл после завершения может повториться или остановиться. Если бы все поперечные

Электромеханическое сопряжение. Трансформация возбуждения в сокращение, по существу представляющая передачу сигнала
Электромеханическое сопряжение. Трансформация возбуждения в сокращение, по существу представляющая передачу сигнала

Остановка сокращения
Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного
Остановка сокращения
Сигналом к остановке сокращения скелетной мышцы служит прекращение нервного

4-3. ВИДЫ И РЕЖИМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ. ОДИНОЧНОЕ СОКРАЩЕНИЕ. ПОНЯТИЕ О ТЕТАНУСЕ.
4-3. ВИДЫ И РЕЖИМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ. ОДИНОЧНОЕ СОКРАЩЕНИЕ. ПОНЯТИЕ О ТЕТАНУСЕ.

Когда мышца укорачивается, преодолевая постоянную нагрузку, говорят об изотоническом сокращении. Большинство
Когда мышца укорачивается, преодолевая постоянную нагрузку, говорят об изотоническом сокращении. Большинство

Одиночное сокращение и тетанус. Одиночный стимул спустя короткое время, около 50
Одиночное сокращение и тетанус. Одиночный стимул спустя короткое время, около 50

Если следующий нервный импульс приходит к волокну раньше, чем закончилось расслабление
Если следующий нервный импульс приходит к волокну раньше, чем закончилось расслабление

При достаточно высокой частоте стимуляции выходящий из ретикулума Са2+ не успевает
При достаточно высокой частоте стимуляции выходящий из ретикулума Са2+ не успевает

Сила сокращения скелетных мышц.
Двигательная единица. Под двигательной единицей понимают каждую
Сила сокращения скелетных мышц.
Двигательная единица. Под двигательной единицей понимают каждую



Максимальная сила мышц.
Абсолютная сила, развиваемая разными мышцами,
неодинакова. Она определяется:
Максимальная сила мышц.
Абсолютная сила, развиваемая разными мышцами,
неодинакова. Она определяется:


4.4. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И СВОЙСТВА гладких мышц.
Классификация гладких мышц. Висцеральные
4.4. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ И СВОЙСТВА гладких мышц.
Классификация гладких мышц. Висцеральные


Иннервация гладких мышц.
Висцеральная гладкая мышца имеет двойную иннервацию — симпатическую
Иннервация гладких мышц.
Висцеральная гладкая мышца имеет двойную иннервацию — симпатическую


Нервно-мышечная передача. Возбуждающее влияние адренергических или холинергических нервов электрически проявляется в
Нервно-мышечная передача. Возбуждающее влияние адренергических или холинергических нервов электрически проявляется в

В периоды состояния относительного покоя величина мембранного потенциала в среднем равна
В периоды состояния относительного покоя величина мембранного потенциала в среднем равна

Автоматия. Многим гладким мышцам свойственна спонтанная, автоматическая активность. Для нее характерно
Автоматия. Многим гладким мышцам свойственна спонтанная, автоматическая активность. Для нее характерно

Механизм сокращения. В механизме сокращения гладкой мышцы имеется особенность, отличающая его
Механизм сокращения. В механизме сокращения гладкой мышцы имеется особенность, отличающая его



Химическая чувствительность. Гладкие мышцы обладают высокой чувствительностью к различным физиологически активным
Химическая чувствительность. Гладкие мышцы обладают высокой чувствительностью к различным физиологически активным

Проведение возбуждения по гладкой мышце. В отличие от того, что имеет
Проведение возбуждения по гладкой мышце. В отличие от того, что имеет

Так же, как и в скелетной мускулатуре, в гладкой потенциалы действия
Так же, как и в скелетной мускулатуре, в гладкой потенциалы действия

Вследствие медленности сокращения гладкая мышца даже при редких ритмических раздражениях (10-12
Вследствие медленности сокращения гладкая мышца даже при редких ритмических раздражениях (10-12
 Геморрагическая лихорадка с почечным синдромом 59
Геморрагическая лихорадка с почечным синдромом 59 Адаптация как процесс вхождения человека в новую для него среду
Адаптация как процесс вхождения человека в новую для него среду Жас кезеңдеріне байланысты баланың жақ-бет аймағы құрылысының ерекшеліктері. Стоматологиялық және соматикалық аурулардың
Жас кезеңдеріне байланысты баланың жақ-бет аймағы құрылысының ерекшеліктері. Стоматологиялық және соматикалық аурулардың Кислородная емкость крови
Кислородная емкость крови Общая характеристика инфекционных заболеваний. Классификация инфекционных болезней, диагностика, лечение, профилактика
Общая характеристика инфекционных заболеваний. Классификация инфекционных болезней, диагностика, лечение, профилактика Адренергичексие средства
Адренергичексие средства Искусственный интеллект в медицине
Искусственный интеллект в медицине Показатели здоровья, их определение и оценка
Показатели здоровья, их определение и оценка Тактильная система
Тактильная система Закономерности нервно-психического развития детей раннего возраста
Закономерности нервно-психического развития детей раннего возраста Хейлопластикаға көрсеткіш,реабилитация, диспансеризация
Хейлопластикаға көрсеткіш,реабилитация, диспансеризация Смерть и процесс умирания. (Лекция 3)
Смерть и процесс умирания. (Лекция 3) Захворювання новонароджених дітей. Захворювання шкіри, пупкової ранки, сепсис
Захворювання новонароджених дітей. Захворювання шкіри, пупкової ранки, сепсис Язвенно-некротический гингивостоматит Венсана
Язвенно-некротический гингивостоматит Венсана Тактика участкового терапевта при заболеваниях щитовидной железы: факторы риска, раннее выявление заболевания
Тактика участкового терапевта при заболеваниях щитовидной железы: факторы риска, раннее выявление заболевания Ісік ауруларының алдын алу кезіндегі рациондарды гигиеналық бағалау әдістемесі
Ісік ауруларының алдын алу кезіндегі рациондарды гигиеналық бағалау әдістемесі Периоды детского возраста
Периоды детского возраста «Школьный» стресс и как с ним бороться
«Школьный» стресс и как с ним бороться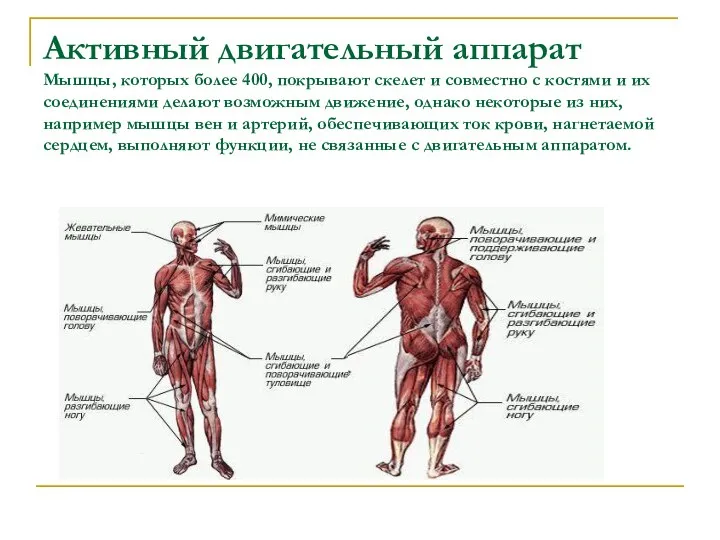 Активный двигательный аппарат
Активный двигательный аппарат Модели сестринского дела. Иерархия потребностей человека
Модели сестринского дела. Иерархия потребностей человека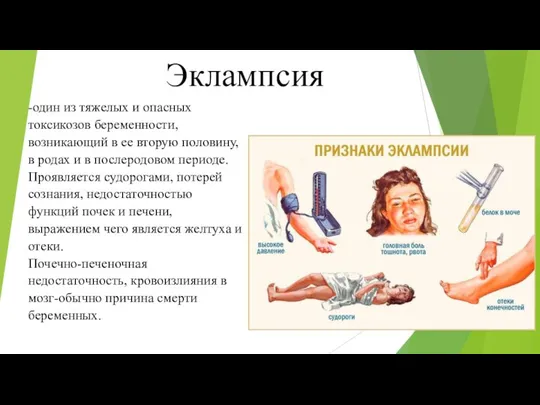 Эклампсия
Эклампсия Свободное и информированное согласие на лечение и участие в эксперименте
Свободное и информированное согласие на лечение и участие в эксперименте Иммунопатологические процессы тканей
Иммунопатологические процессы тканей Водно-солевой обмен
Водно-солевой обмен Болезни кожи
Болезни кожи Воспалительные и дистрофические заболевания ВНЧС у детей
Воспалительные и дистрофические заболевания ВНЧС у детей Адамның тамақтануында тропикалық жемістердің маңызы
Адамның тамақтануында тропикалық жемістердің маңызы Лучевое исследование лёгких, плевры
Лучевое исследование лёгких, плевры