- Т-клеточный антигенспецифический приобретённый иммунитет

Содержание
- 2. Лимфоциты – основные клетки иммунной системы, которые распознают чужеродные антигены и осуществляют реакции, приводящие к удалению
- 4. Характеристика основных популяций лимфоцитов
- 5. 2-х этапная дифференцировка лимфоцитов: I этап – Аг-независимый ( в костном мозге или тимусе), лимфопоэз (происходит
- 6. В органы лимфопоэза «заходят» недифференцированные клетки-предшественники, из которых в костном мозге (для В лимфоцитов) или тимусе
- 7. Популяция лимфоцитов имеет клональную структуру. Клон лимфоцитов - тождественные друг другу клетки, поделившиеся правильным митозом и
- 8. Т лимфоциты - thymus-dependent играют центральную роль в развитии адаптивного (приобретенного) иммунного ответа и представляют собой
- 9. Имеется 2 типа TCR: αβTCR– 80% γδ TCR – 20% Каждая функционально зрелая Т клетка имеет
- 10. Антитела (иммуноглобулины) – это белки, которые синтезируются под влиянием антигенов и специфически с ними реагируют Fc
- 11. Основные отличия TCR от BCR: TCR – моновалентен Существует только в виде поверхностного рецептора и не
- 12. CD3 комплекс включает 5 белков: 3 белка γ, δ, ε и димер ζζ. К TCR имеют
- 13. Сравнение αβ и γδ Т клеток
- 14. Основные популяции Т лимфоцитов
- 15. Функциональные субпопуляции Т хелперов (T helper) 1.Клеточный иммунитет 2. Образование CTL 3. Активация макрофагов 4. Иммунное
- 16. Функциональные субклассы CD4 Т лимфоцитов Tfh
- 17. ДИФФЕРЕНЦИРОВКА Т-ХЕЛПЕРНЫХ КЛОНОВ Тх0 Тх1 Тх2 Tx17 Tx10 ? IL-4 IL-23 TSLP IFN-γ IL-2 TNF IL-4
- 18. C – cluster D – differentiation (definition) АНТИГЕНЫ – выявляются с помощью моноклональных антител CD АНТИГЕНЫ
- 19. Т-лимфоциты Панель моноклональных антител: CD3 – Т лимфоциты CD4 - Т-хелперы CD8 - Т- цитотоксические/эффекторы HLA
- 20. Основная функция зрелых Т лимфоцитов – распознавание чужеродных антигенных пептидов в комплексе с собственными Аг МНС
- 21. Дифференцировка и селекция Т лимфоцитов в тимусе Т лимфоциты являются единственным типом клеток крови, для которых
- 22. Дифференцировка и селекция Т лимфоцитов в тимусе I этап – формирование TCR при контакте с клетками-’кормилицами’
- 23. Дифференцировка и селекция Т лимфоцитов в тимусе II этап – «позитивная» селекция - клональная пролиферация –
- 24. Дифференцировка и селекция Т лимфоцитов в тимусе III этап – «отрицательная» селекция. Клональная делеция происходит в
- 25. В результате «позитивной и «негативной» селекции из тимуса в кровоток и лимфоидные органы поступают только такие
- 26. Миграция и заселение периферии Зрелые Т лимфоциты покидают тимус через сосуды кортико-медулярной зоны, 2-5%, т.е. 1х106
- 27. Миграция и заселение периферии Взаимодействие между L-селектинами и адгезивными молекулами эндотелиальных клеток лежит в основе специфического
- 28. Итак, Основной источник клеток иммунной системы – костный мозг Т лимфоциты развиваются в тимусе. Здесь формируется
- 29. Т-клеточный ответ В основе реакций иммунной системы лежит распознавание чужеродный антигенов специфическими клонами лимфоцитов с последующей
- 30. T лимфоциты не распознают нативные антигены Y Y Y Y Y Y Пролиферация и продукция антител
- 31. Т лимфоцит распознает переработанный антиген T cell response No T cell response No T cell response
- 32. Этапы Т-клеточного ответа: I - Начальный процессинг презентация II – Активация Т лимфоцитов III – Формирование
- 33. Клеточный иммунный ответ инициируется антигеном, который проникает внутрь антиген-презентирующей клетки (АПК, АРС – англ.)! АПК: ДК
- 34. Процессинг антигена (переработка) заключается в его расщеплении на пептидные фрагменты. В зависимости от Аг процессинг происходит
- 35. При этом, Т-клетки, рестриктированные в отношении распознавания антигена молекулами МНС I класса (CD8+ Т лимфоциты), распознают
- 36. Две цепи молекулы МНС образуют т.н. «расщелину», способную связывать после внутриклеточного ферментативного расщепления фрагменты Аг и
- 37. Процессинг I типа Протеасомы расщепляют белки и высвобождают пептиды в цитоплазму. В систему процессинга включаются белки
- 38. ENDOPLASMIC RETICULUM CYTOSOL Пептиды антигенов образуются в цитоплазме отдельно от образующихся MHC I класса в ER
- 39. Transporters associated with antigen processing (TAP1 & 2) Транспортер переносит пептиды размером 8-12 аминокислот, т.е. Аг
- 40. Молекула МНС приобретает стабильную форму только после связывания с пептидом. После этого молекула готова мигрировать на
- 42. Процессинг II типа осуществляется по отношению к экзогенным белкам в специализированных клетках (преимущественно в Мн/Мф). Антигенные
- 43. Y Пиноцитоз Фагоцитоз Ig-опосредованный фагоцитоз Поглощение экзогенных антигенов СR-опосредованный фагоцитоз (опсонизация) Fc R-опосредованный фагоцитоз (опсонизация) После
- 44. Протеазы расщепляют белок до пептидов размером ~24 аминокислот Экзогенный процессинг (II) Антигенные пептиды в эндосоме Катепсин
- 45. Молекулы МНС II класса синтезируются в ER, откуда после их временного соединения с инвариантной цепью (Ii)
- 46. Функции: Стабилизация МНС II в ER Предотвращает преждевременное связывание антигенного пептида с MHC II Инвариантная цепь
- 47. Class II associated invariant chain peptide (CLIP) (αβinv)3 комплексы направляются в эндосомы Катепсин L деградирует инвариантную
- 48. Цитоплазма Мф Процессинг II типа
- 49. T и B клетки распознают антиген различными способами Для распознавания Т клетками антиген должен быть переработан/процессирован
- 50. Презентация антигена Представление (экспрессия) антигена на поверхности (антиген-презентирующих) клеток в форме, доступной для распознавания лимфоцитами, т.е.
- 51. αβ TCR в комплексе с мембранной молекулой CD4 или CD8 способен связывать (т.е. распознавать ) только
- 52. Презентация АГ CD4 T cell CD8 CTL
- 53. За открытие того факта, что Т лимфоциты способны распознавать только пептидные антигены и только при условии,
- 54. В процессе иммунного ответа на Аг образуется тримолекулярный комплекс, состоящий из Аг, МНС и TCR Комплексы
- 55. CD4/8 coreceptors CD4 и CD8 молекулы действуют как “кo-рецепторы”: они распознают MHC II или MHC I
- 56. CD28 provides 2nd signal Ко-стимулирующие молекулы усиливают активацию Т лимфоцитов
- 57. Поверхностные молекулы антиген-презентирующих клеток MHC молекулы:Class I, Class II Костимулирующие молекулы:B7(B7-1(CD80)、B7-2(CD86)) Адгезивные молекулы:CD58(LFA-3), CD54(intercellular adhesion molecule-1,ICAM-1)
- 58. Механизмы презентации Т-хелпер АПК ЦТЛ Клетка-мишень
- 59. После связывания рецепторами соответствующих лигандов запускаются цепи молекулярных событий, осуществляющие внутриклеточную передачу сигналов от рецепторов к
- 60. Поскольку активация служит подготовкой к делению клеток, ее основной результат состоит в индукции ряда генов, продукты
- 61. Процесс активации кодируется 2-х сигнальной системой: В результате связывания комплекса антиген (пептида)+МНС с ТСR – основной
- 62. Активация Т лимфоцитов Scharenberg et al. Nat Rev Immunol. 2007
- 63. Lck: - семейство Src тирозинкиназ, которые нековалентно связаны с цитоплазматическими доменами CD4 и CD8 молекул. Lck
- 64. LAT
- 65. ZAP-70 (Zeta-associated protein of 70 kD): связывается с фосфорилированными тирозинами в цитоплазматических доменах CD3 и ζ
- 66. LAT
- 67. Phospholipase Cγ (PLCγ): фермент, который гидролизирует активные компоненты плазматической мембраны (фосфоинозитиды), в результате чего генерируются вторичные
- 68. Выделяют несколько параллельных взаимодополняющих механизмов формирования и проведения активационных сигналов в Т-лимфоцитах. Путь активации кальциневрина и
- 69. LAT
- 70. Начиная с образования IP3 и DAG формируются завершающие звенья реакций, приводящие к формированию транскрипционных факторов NF-AT
- 72. Men F B
- 73. Экспрессия генов в Т лимфоцитах
- 74. Экспрессия генов в Т лимфоцитах
- 75. Экспрессия генов в Т лимфоцитах
- 76. Пролиферация и дифференцировка Т лимфоцитов Это митотическое деление Т лимфоцитов, когда из одной клетки появляются 2
- 77. IL2-R, c-jun, c-fos, c-myc Передача сигнала от цитокиновых рецепторов Рецепция активированными клетками цитокинов генерирует сигналы, необходимые
- 78. ДифференцировкаТ хелперов Клеточный иммунный ответ Гуморальный иммунный ответ IFNγ, IL12 IL 4, 10 TNFβ IL2
- 79. Факторы дифференцировки Т хелперов: Доза Аг Микроокружение органов, в которых происходит дифференцировка Т хелперов Участие АПК
- 81. Скачать презентацию

Лимфоциты – основные клетки иммунной системы,
которые распознают чужеродные антигены и осуществляют
реакции,
Лимфоциты – основные клетки иммунной системы,
которые распознают чужеродные антигены и осуществляют
реакции,
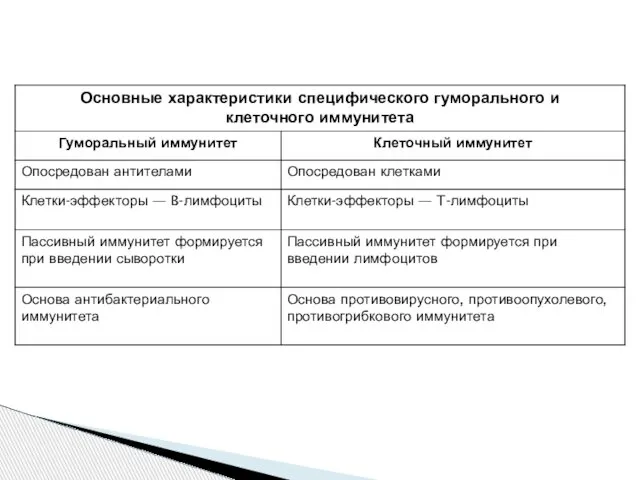
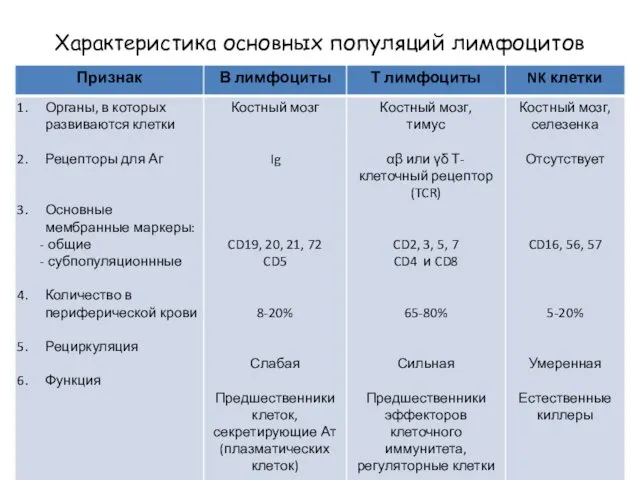
Характеристика основных популяций лимфоцитов
Характеристика основных популяций лимфоцитов

2-х этапная дифференцировка лимфоцитов:
I этап – Аг-независимый ( в костном мозге
2-х этапная дифференцировка лимфоцитов:
I этап – Аг-независимый ( в костном мозге

В органы лимфопоэза «заходят» недифференцированные клетки-предшественники, из которых в костном мозге
В органы лимфопоэза «заходят» недифференцированные клетки-предшественники, из которых в костном мозге

Популяция лимфоцитов имеет клональную
структуру.
Клон лимфоцитов - тождественные друг другу клетки, поделившиеся
Популяция лимфоцитов имеет клональную
структуру.
Клон лимфоцитов - тождественные друг другу клетки, поделившиеся

Т лимфоциты - thymus-dependent
играют центральную роль в развитии адаптивного
(приобретенного) иммунного ответа
Т лимфоциты - thymus-dependent
играют центральную роль в развитии адаптивного
(приобретенного) иммунного ответа
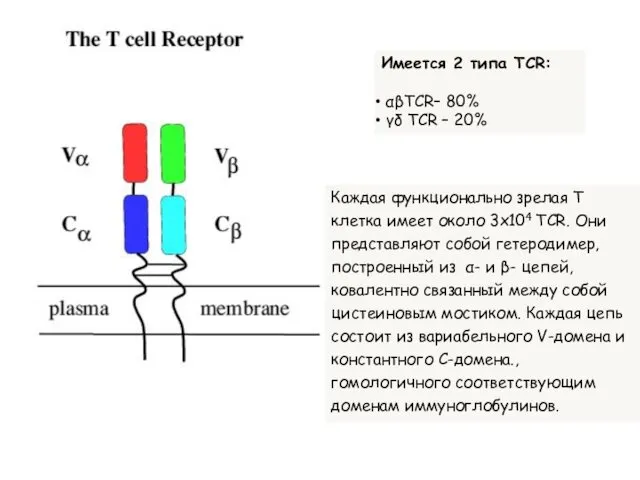
Имеется 2 типа TCR:
αβTCR– 80%
γδ TCR – 20%
Каждая функционально
Имеется 2 типа TCR:
αβTCR– 80%
γδ TCR – 20%
Каждая функционально
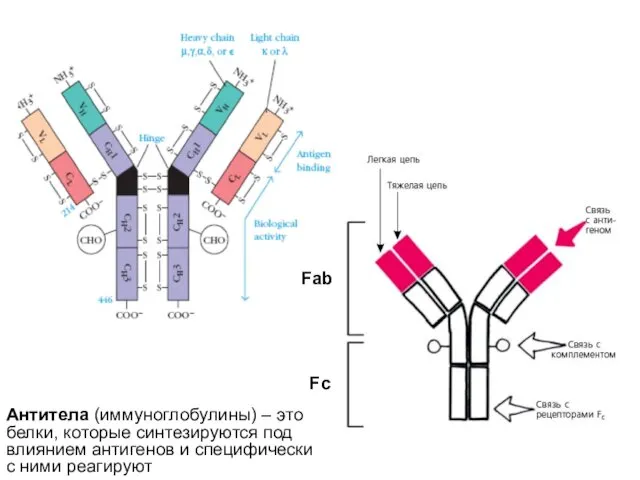
Антитела (иммуноглобулины) – это белки, которые синтезируются под влиянием антигенов и
Антитела (иммуноглобулины) – это белки, которые синтезируются под влиянием антигенов и
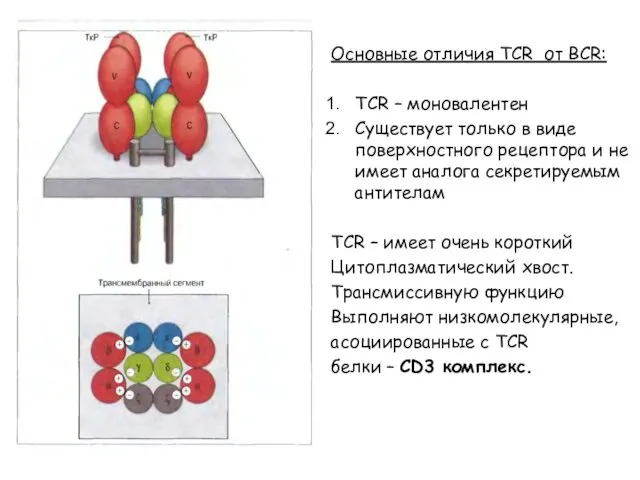
Основные отличия TCR от BCR:
TCR – моновалентен
Существует только в виде поверхностного
Основные отличия TCR от BCR:
TCR – моновалентен
Существует только в виде поверхностного
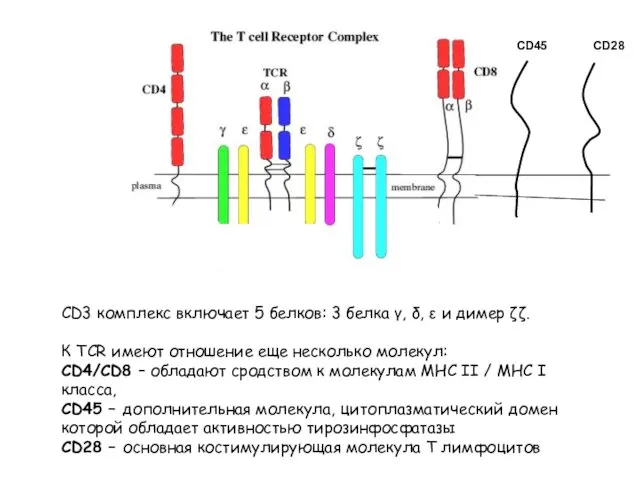
CD3 комплекс включает 5 белков: 3 белка γ, δ, ε и
CD3 комплекс включает 5 белков: 3 белка γ, δ, ε и
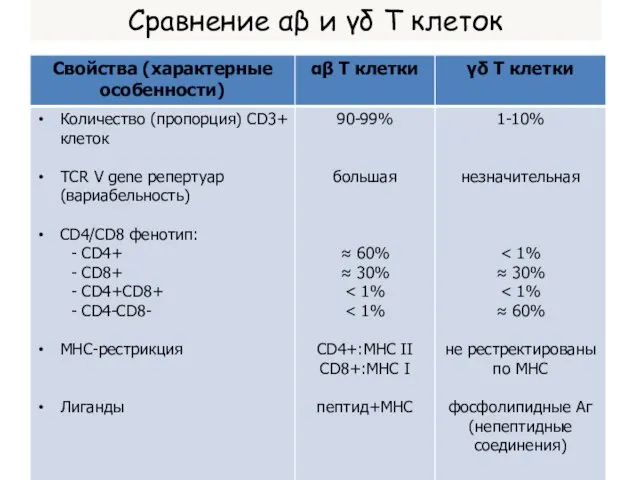
Сравнение αβ и γδ Т клеток
Сравнение αβ и γδ Т клеток
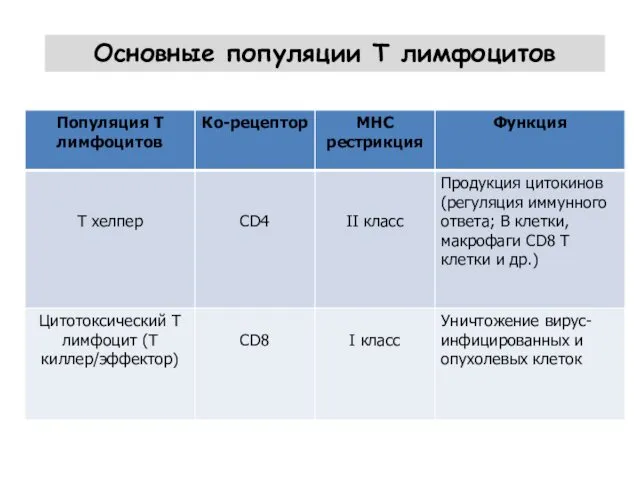
Основные популяции Т лимфоцитов
Основные популяции Т лимфоцитов
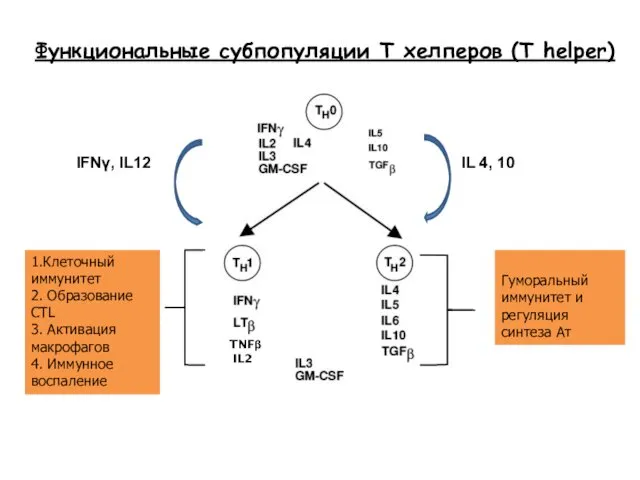
Функциональные субпопуляции Т хелперов (T helper)
1.Клеточный иммунитет
2. Образование CTL
3. Активация макрофагов
4.
Функциональные субпопуляции Т хелперов (T helper)
1.Клеточный иммунитет
2. Образование CTL
3. Активация макрофагов
4.
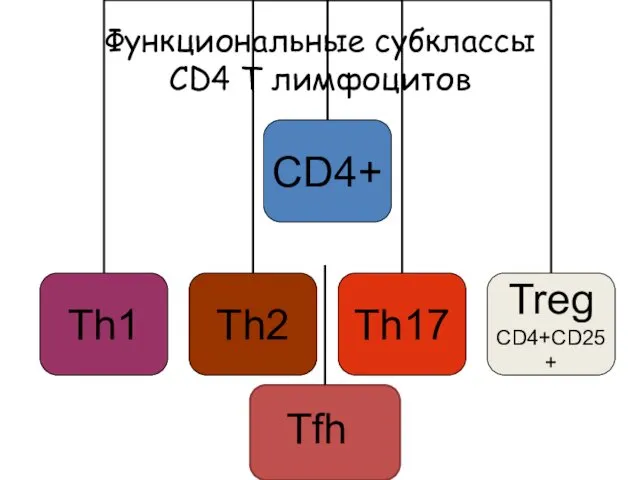
Функциональные субклассы
CD4 Т лимфоцитов
Tfh
Функциональные субклассы
CD4 Т лимфоцитов
Tfh
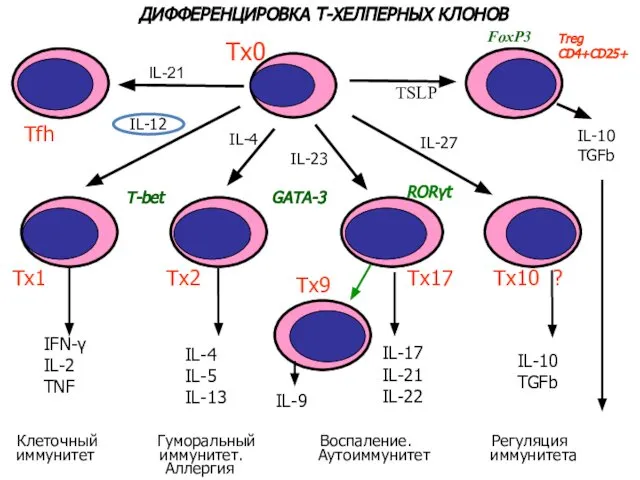
ДИФФЕРЕНЦИРОВКА Т-ХЕЛПЕРНЫХ КЛОНОВ
Тх0
Тх1 Тх2 Tx17 Tx10 ?
IL-4
ДИФФЕРЕНЦИРОВКА Т-ХЕЛПЕРНЫХ КЛОНОВ
Тх0
Тх1 Тх2 Tx17 Tx10 ?
IL-4

C – cluster
D – differentiation (definition)
АНТИГЕНЫ – выявляются с помощью
моноклональных
C – cluster
D – differentiation (definition)
АНТИГЕНЫ – выявляются с помощью моноклональных
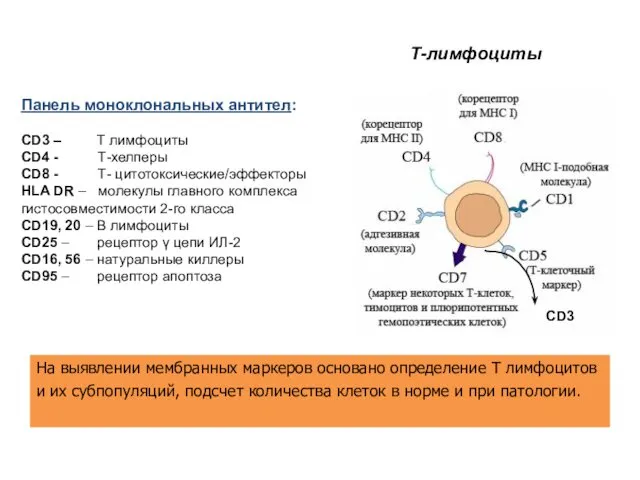
Т-лимфоциты
Панель моноклональных антител:
CD3 – Т лимфоциты
CD4 - Т-хелперы
CD8 - Т-
Т-лимфоциты
Панель моноклональных антител:
CD3 – Т лимфоциты
CD4 - Т-хелперы
CD8 - Т-

Основная функция зрелых Т лимфоцитов – распознавание чужеродных антигенных пептидов в
Основная функция зрелых Т лимфоцитов – распознавание чужеродных антигенных пептидов в

Дифференцировка и селекция Т лимфоцитов в тимусе
Т лимфоциты являются единственным типом
Дифференцировка и селекция Т лимфоцитов в тимусе
Т лимфоциты являются единственным типом
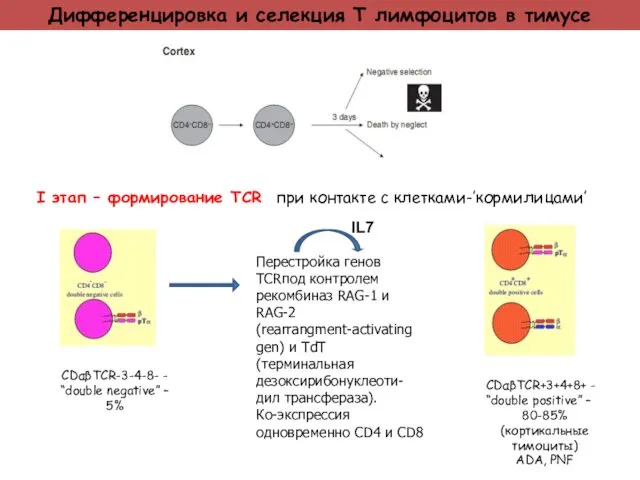
Дифференцировка и селекция Т лимфоцитов в тимусе
I этап – формирование TCR
Дифференцировка и селекция Т лимфоцитов в тимусе
I этап – формирование TCR

Дифференцировка и селекция Т лимфоцитов в тимусе
II этап – «позитивная» селекция
Дифференцировка и селекция Т лимфоцитов в тимусе
II этап – «позитивная» селекция
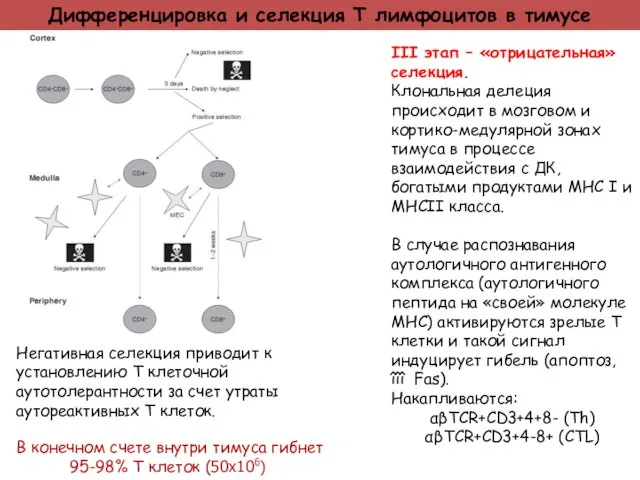
Дифференцировка и селекция Т лимфоцитов в тимусе
III этап – «отрицательная» селекция.
Клональная
Дифференцировка и селекция Т лимфоцитов в тимусе
III этап – «отрицательная» селекция.
Клональная

В результате «позитивной и «негативной» селекции из тимуса в кровоток и
В результате «позитивной и «негативной» селекции из тимуса в кровоток и

Миграция и заселение периферии
Зрелые Т лимфоциты покидают тимус через сосуды
Миграция и заселение периферии
Зрелые Т лимфоциты покидают тимус через сосуды
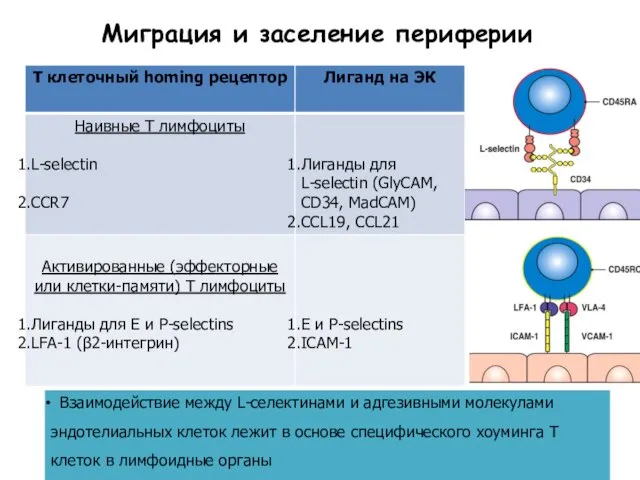
Миграция и заселение периферии
Взаимодействие между L-селектинами и адгезивными молекулами эндотелиальных
Миграция и заселение периферии
Взаимодействие между L-селектинами и адгезивными молекулами эндотелиальных

Итак,
Основной источник клеток иммунной системы – костный мозг
Т лимфоциты развиваются в
Итак,
Основной источник клеток иммунной системы – костный мозг
Т лимфоциты развиваются в

Т-клеточный ответ
В основе реакций иммунной системы лежит распознавание чужеродный антигенов специфическими
Т-клеточный ответ
В основе реакций иммунной системы лежит распознавание чужеродный антигенов специфическими
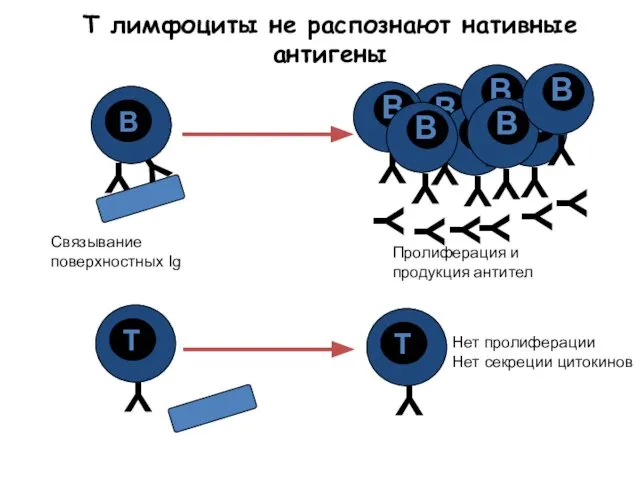
T лимфоциты не распознают нативные антигены
Y
Y
Y
Y
Y
Y
Пролиферация и продукция антител
Нет пролиферации
Нет секреции
T лимфоциты не распознают нативные антигены
Y
Y
Y
Y
Y
Y
Пролиферация и продукция антител
Нет пролиферации
Нет секреции
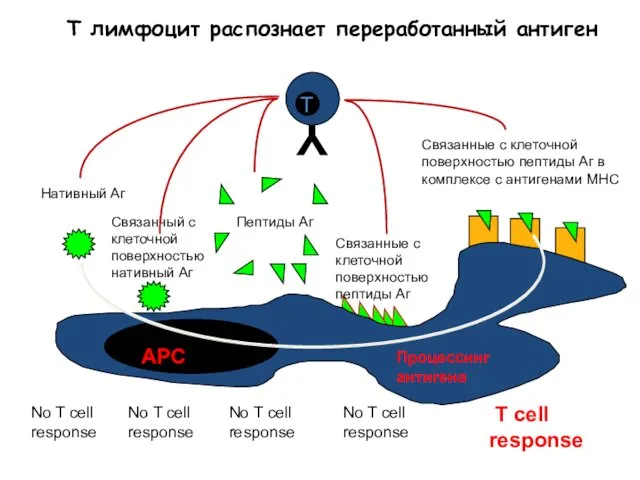
Т лимфоцит распознает переработанный антиген
T cell
response
No T cell
response
No T cell
response
No
Т лимфоцит распознает переработанный антиген
T cell
response
No T cell
response
No T cell
response
No
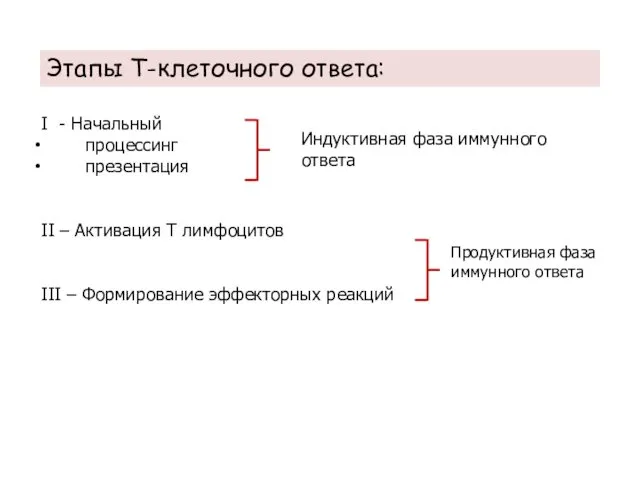
Этапы Т-клеточного ответа:
I - Начальный
процессинг
презентация
II – Активация Т лимфоцитов
III
Этапы Т-клеточного ответа:
I - Начальный
процессинг
презентация
II – Активация Т лимфоцитов
III

Клеточный иммунный ответ инициируется антигеном, который проникает внутрь антиген-презентирующей клетки (АПК,
Клеточный иммунный ответ инициируется антигеном, который проникает внутрь антиген-презентирующей клетки (АПК,

Процессинг антигена (переработка) заключается в его расщеплении на пептидные фрагменты.
В зависимости
В зависимости

При этом,
Т-клетки, рестриктированные в отношении распознавания антигена молекулами МНС I
При этом,
Т-клетки, рестриктированные в отношении распознавания антигена молекулами МНС I
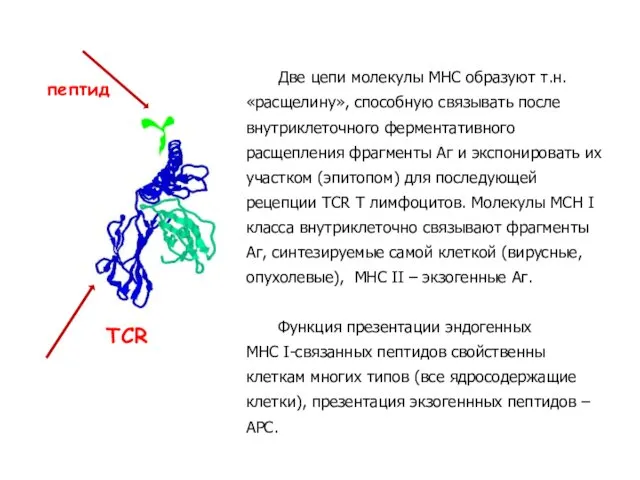
Две цепи молекулы МНС образуют т.н. «расщелину», способную связывать после внутриклеточного
Две цепи молекулы МНС образуют т.н. «расщелину», способную связывать после внутриклеточного
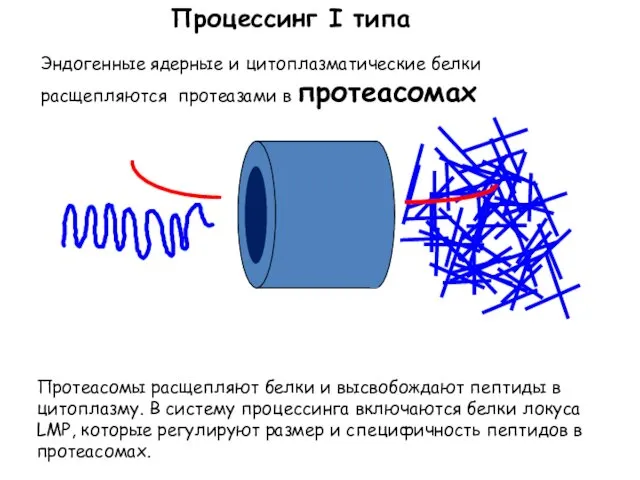
Процессинг I типа
Протеасомы расщепляют белки и высвобождают пептиды в цитоплазму. В
Процессинг I типа
Протеасомы расщепляют белки и высвобождают пептиды в цитоплазму. В
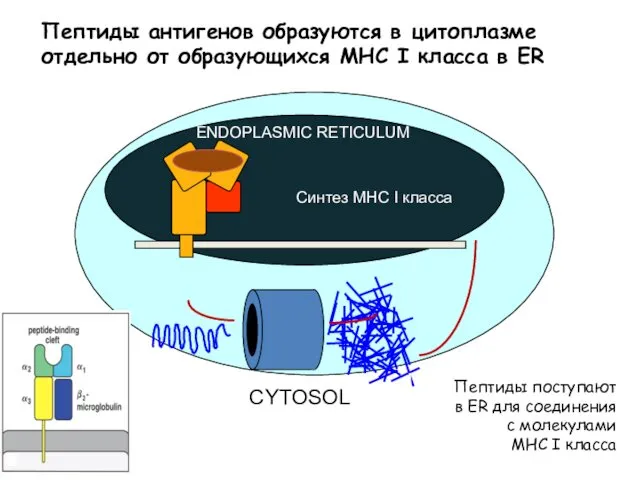
ENDOPLASMIC RETICULUM
CYTOSOL
Пептиды антигенов образуются в цитоплазме отдельно от образующихся MHC I
ENDOPLASMIC RETICULUM
CYTOSOL
Пептиды антигенов образуются в цитоплазме отдельно от образующихся MHC I
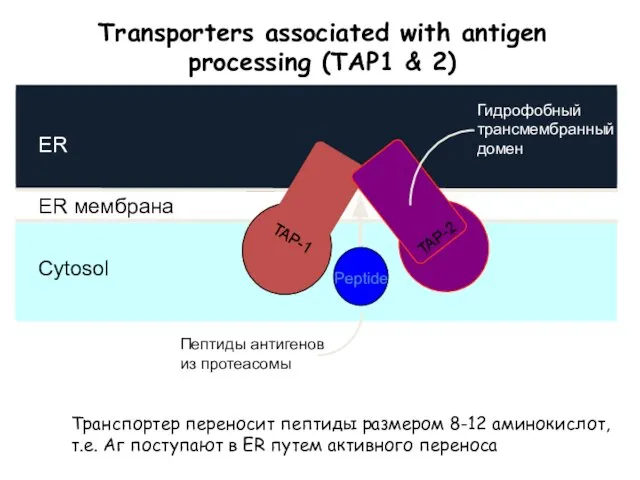
Transporters associated with antigen processing (TAP1 & 2)
Транспортер переносит пептиды размером
Transporters associated with antigen processing (TAP1 & 2)
Транспортер переносит пептиды размером

Молекула МНС приобретает стабильную форму только после связывания с пептидом. После
Молекула МНС приобретает стабильную форму только после связывания с пептидом. После


Процессинг II типа
осуществляется по отношению к экзогенным белкам в специализированных клетках
Процессинг II типа
осуществляется по отношению к экзогенным белкам в специализированных клетках
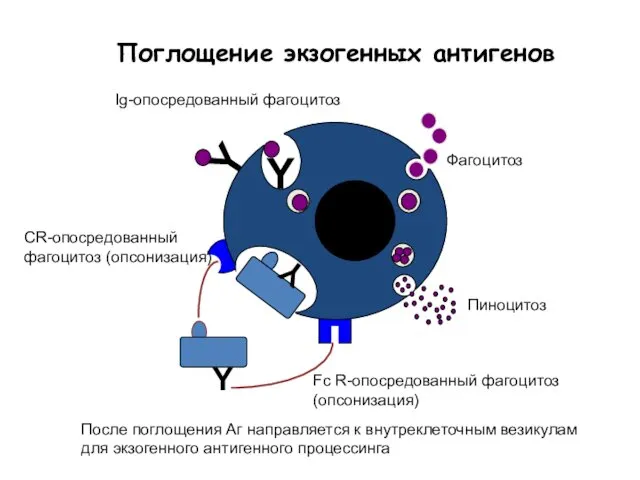
Y
Пиноцитоз
Фагоцитоз
Ig-опосредованный фагоцитоз
Поглощение экзогенных антигенов
СR-опосредованный
фагоцитоз (опсонизация)
Fc R-опосредованный фагоцитоз (опсонизация)
После поглощения Аг
Y
Пиноцитоз
Фагоцитоз
Ig-опосредованный фагоцитоз
Поглощение экзогенных антигенов
СR-опосредованный
фагоцитоз (опсонизация)
Fc R-опосредованный фагоцитоз (опсонизация)
После поглощения Аг
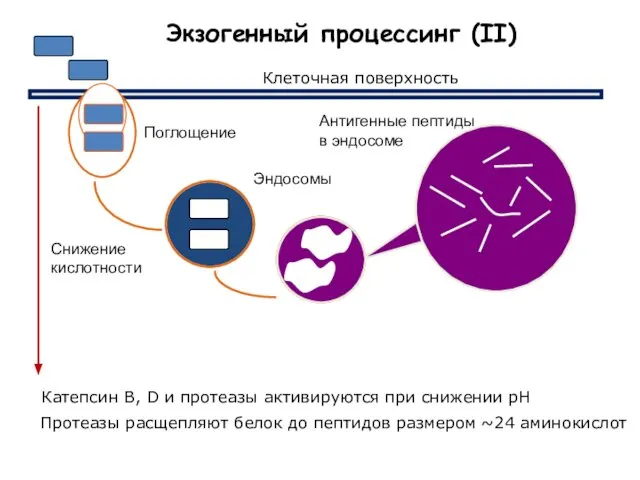
Протеазы расщепляют белок до пептидов размером ~24 аминокислот
Экзогенный процессинг (II)
Антигенные пептиды
Протеазы расщепляют белок до пептидов размером ~24 аминокислот
Экзогенный процессинг (II)
Антигенные пептиды

Молекулы МНС II класса синтезируются в ER, откуда после их временного
Молекулы МНС II класса синтезируются в ER, откуда после их временного
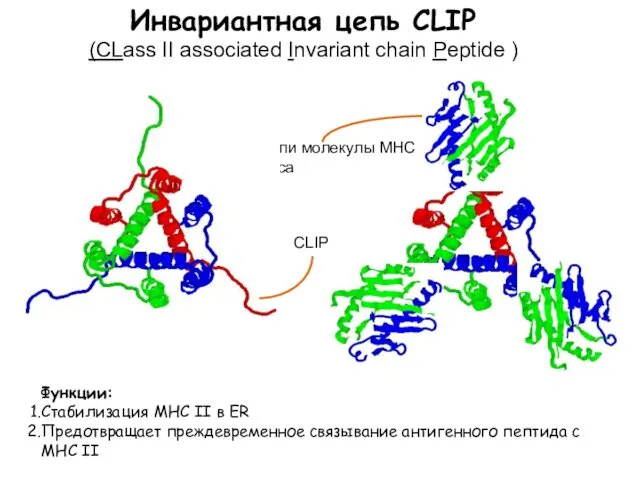
Функции:
Стабилизация МНС II в ER
Предотвращает преждевременное связывание антигенного пептида с MHC
Функции:
Стабилизация МНС II в ER
Предотвращает преждевременное связывание антигенного пептида с MHC
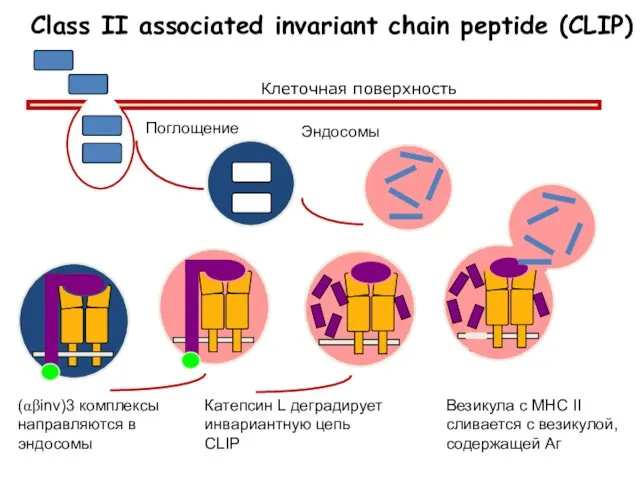
Class II associated invariant chain peptide (CLIP)
(αβinv)3 комплексы
направляются в
эндосомы
Катепсин L
Class II associated invariant chain peptide (CLIP)
(αβinv)3 комплексы
направляются в
эндосомы
Катепсин L
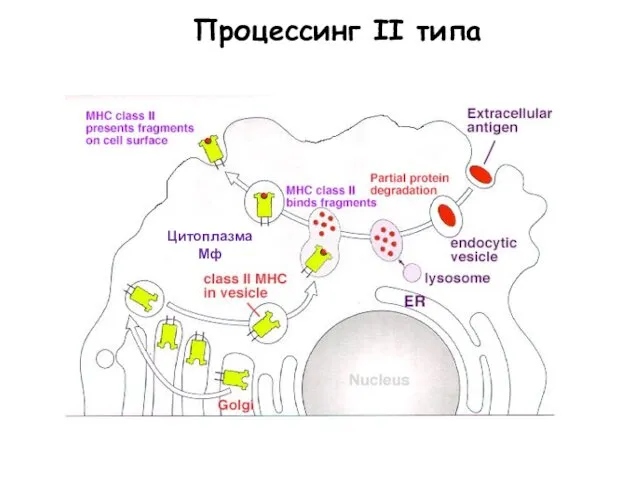
Цитоплазма Мф
Процессинг II типа
Цитоплазма Мф
Процессинг II типа

T и B клетки распознают антиген различными способами
Для распознавания Т клетками
T и B клетки распознают антиген различными способами
Для распознавания Т клетками

Презентация антигена
Представление (экспрессия) антигена на поверхности (антиген-презентирующих) клеток в форме, доступной
Презентация антигена
Представление (экспрессия) антигена на поверхности (антиген-презентирующих) клеток в форме, доступной

αβ TCR в комплексе с мембранной молекулой CD4 или CD8 способен
αβ TCR в комплексе с мембранной молекулой CD4 или CD8 способен
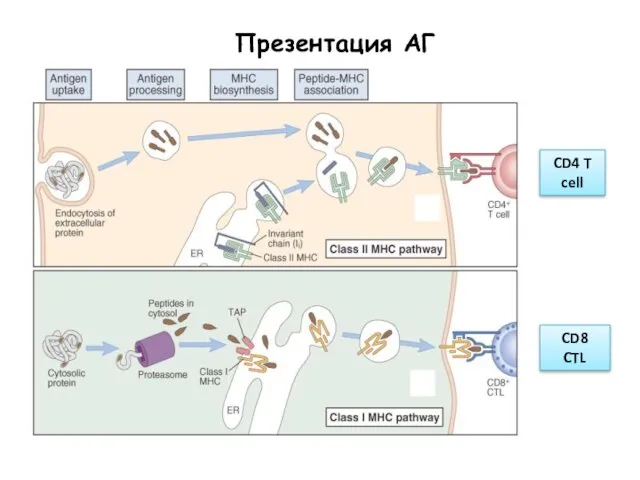
Презентация АГ
CD4 T cell
CD8
CTL
Презентация АГ
CD4 T cell
CD8
CTL
За открытие того факта, что Т лимфоциты способны распознавать только пептидные
За открытие того факта, что Т лимфоциты способны распознавать только пептидные

В процессе иммунного ответа на Аг образуется тримолекулярный комплекс, состоящий из
В процессе иммунного ответа на Аг образуется тримолекулярный комплекс, состоящий из
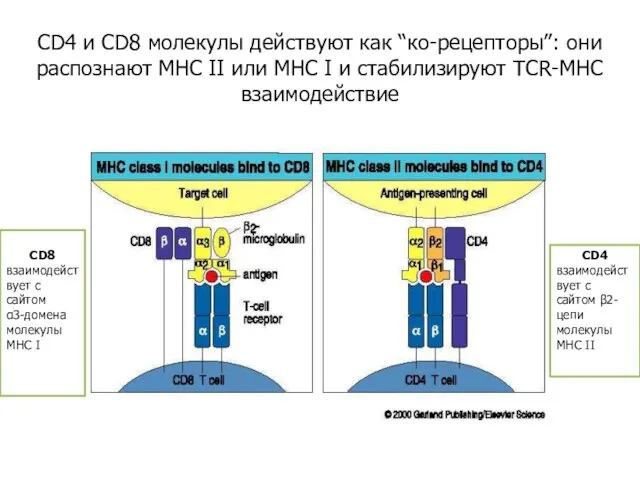
CD4/8 coreceptors
CD4 и CD8 молекулы действуют как “кo-рецепторы”: они распознают MHC
CD4/8 coreceptors
CD4 и CD8 молекулы действуют как “кo-рецепторы”: они распознают MHC
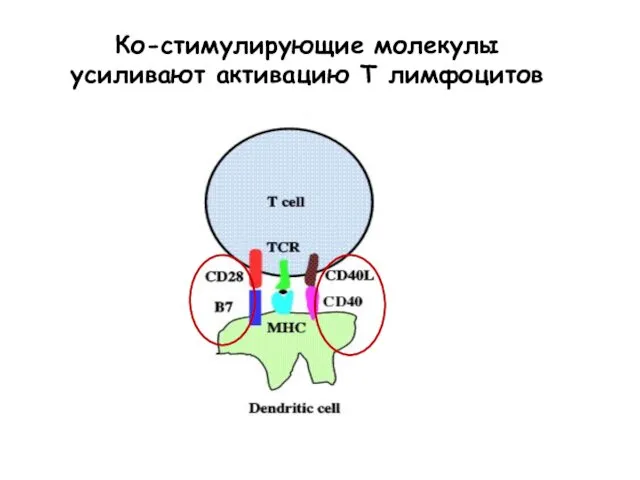
CD28 provides 2nd signal
Ко-стимулирующие молекулы усиливают активацию Т лимфоцитов
CD28 provides 2nd signal
Ко-стимулирующие молекулы усиливают активацию Т лимфоцитов

Поверхностные молекулы антиген-презентирующих клеток
MHC молекулы:Class I, Class II
Костимулирующие молекулы:B7(B7-1(CD80)、B7-2(CD86))
Адгезивные молекулы:CD58(LFA-3), CD54(intercellular
Поверхностные молекулы антиген-презентирующих клеток
MHC молекулы:Class I, Class II
Костимулирующие молекулы:B7(B7-1(CD80)、B7-2(CD86))
Адгезивные молекулы:CD58(LFA-3), CD54(intercellular
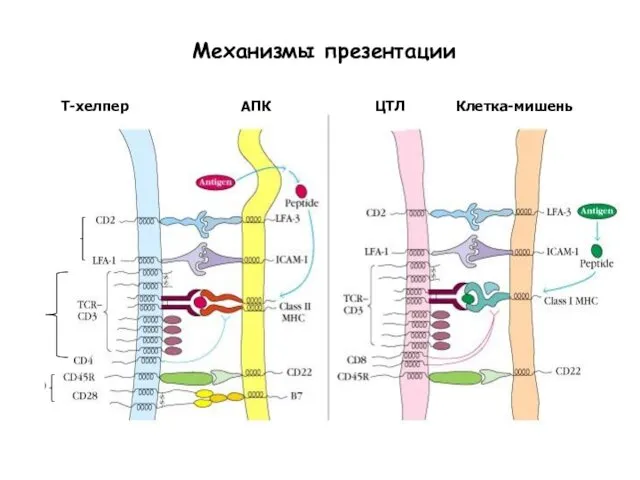
Механизмы презентации
Т-хелпер АПК ЦТЛ Клетка-мишень
Механизмы презентации
Т-хелпер АПК ЦТЛ Клетка-мишень

После связывания рецепторами соответствующих лигандов запускаются цепи молекулярных событий, осуществляющие внутриклеточную
После связывания рецепторами соответствующих лигандов запускаются цепи молекулярных событий, осуществляющие внутриклеточную

Поскольку активация служит подготовкой к делению клеток, ее основной результат состоит
Поскольку активация служит подготовкой к делению клеток, ее основной результат состоит

Процесс активации кодируется 2-х сигнальной системой:
В результате связывания комплекса антиген (пептида)+МНС
Процесс активации кодируется 2-х сигнальной системой:
В результате связывания комплекса антиген (пептида)+МНС

Активация Т лимфоцитов
Scharenberg et al. Nat Rev Immunol. 2007
Активация Т лимфоцитов
Scharenberg et al. Nat Rev Immunol. 2007

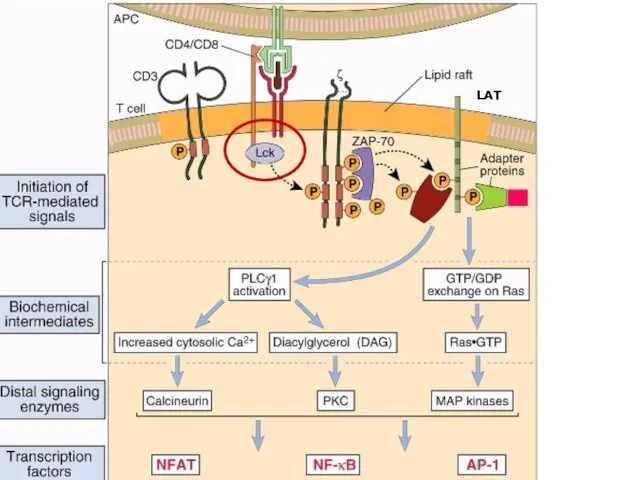
Lck: - семейство Src тирозинкиназ, которые нековалентно связаны с цитоплазматическими доменами
Lck: - семейство Src тирозинкиназ, которые нековалентно связаны с цитоплазматическими доменами
LAT
LAT

ZAP-70 (Zeta-associated protein of 70 kD): связывается с фосфорилированными тирозинами в
ZAP-70 (Zeta-associated protein of 70 kD): связывается с фосфорилированными тирозинами в
LAT
LAT

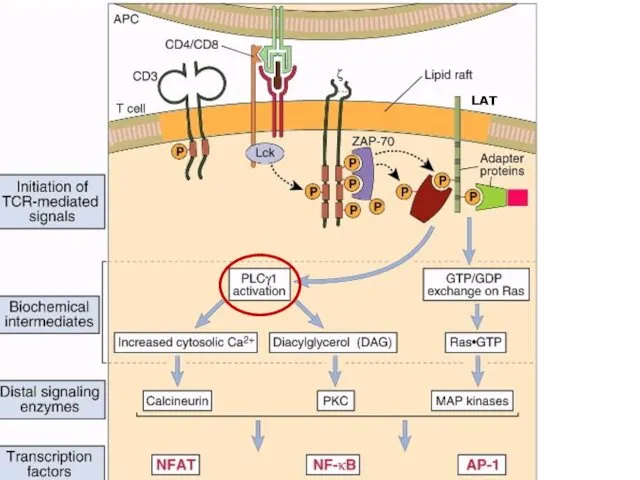
Phospholipase Cγ (PLCγ): фермент, который гидролизирует активные компоненты плазматической мембраны (фосфоинозитиды),
Phospholipase Cγ (PLCγ): фермент, который гидролизирует активные компоненты плазматической мембраны (фосфоинозитиды),

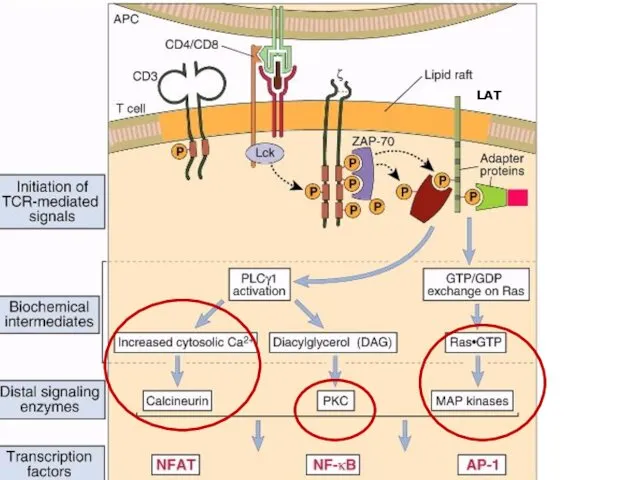
Выделяют несколько параллельных взаимодополняющих механизмов формирования и проведения активационных сигналов в
Выделяют несколько параллельных взаимодополняющих механизмов формирования и проведения активационных сигналов в
LAT
LAT
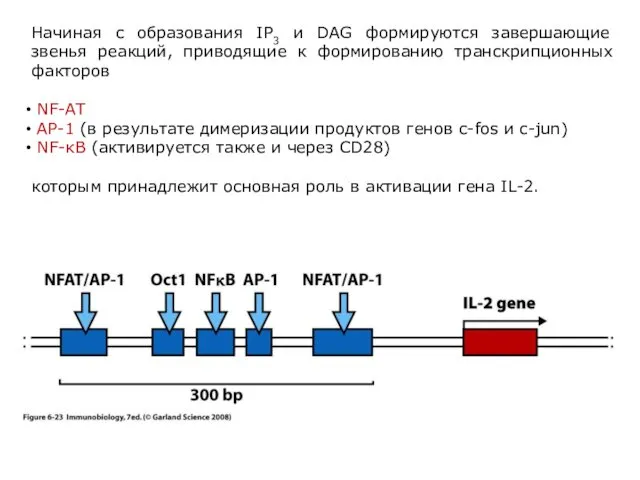
Начиная с образования IP3 и DAG формируются завершающие звенья реакций, приводящие
Начиная с образования IP3 и DAG формируются завершающие звенья реакций, приводящие

Men
F
B
Men
F
B
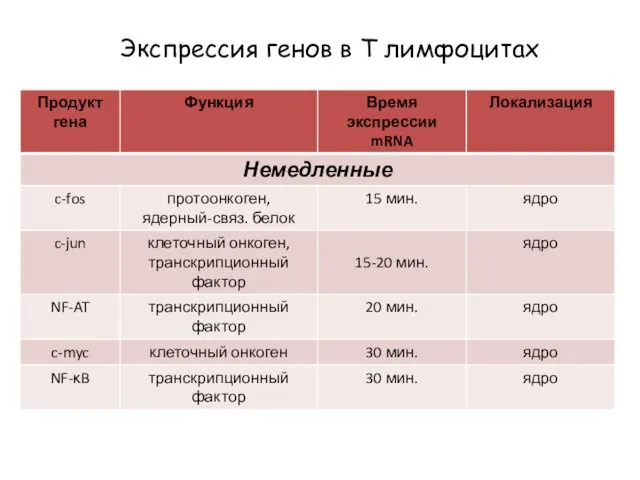
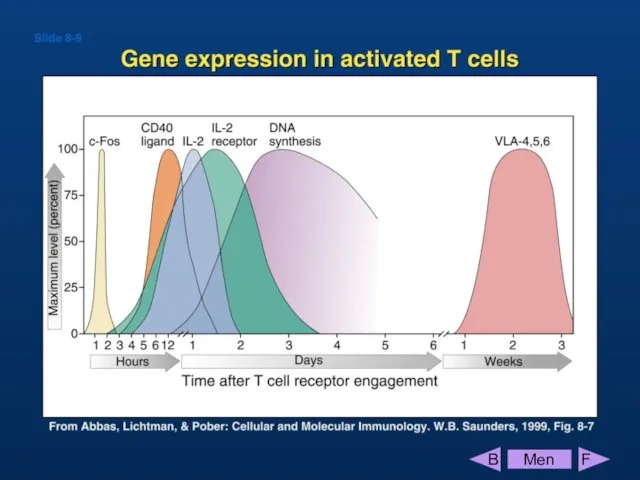
Экспрессия генов в Т лимфоцитах
Экспрессия генов в Т лимфоцитах
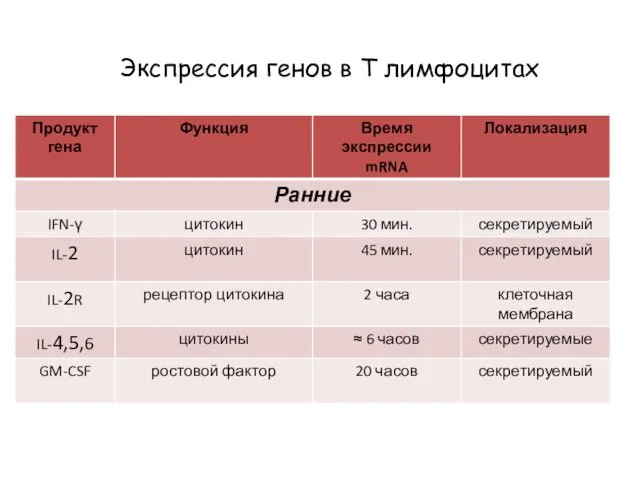
Экспрессия генов в Т лимфоцитах
Экспрессия генов в Т лимфоцитах
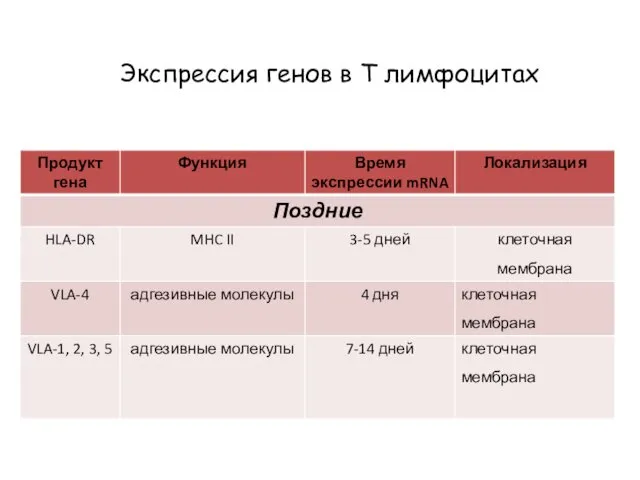
Экспрессия генов в Т лимфоцитах
Экспрессия генов в Т лимфоцитах

Пролиферация и дифференцировка
Т лимфоцитов
Это митотическое деление Т лимфоцитов, когда из
Пролиферация и дифференцировка
Т лимфоцитов
Это митотическое деление Т лимфоцитов, когда из
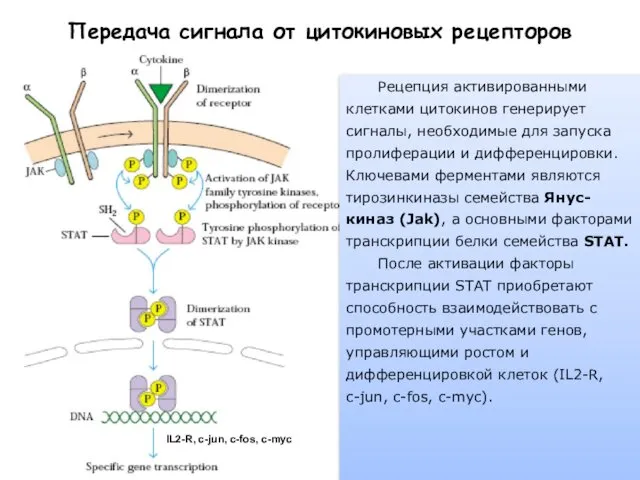
IL2-R, c-jun, c-fos, c-myc
Передача сигнала от цитокиновых рецепторов
Рецепция активированными клетками цитокинов
IL2-R, c-jun, c-fos, c-myc
Передача сигнала от цитокиновых рецепторов
Рецепция активированными клетками цитокинов
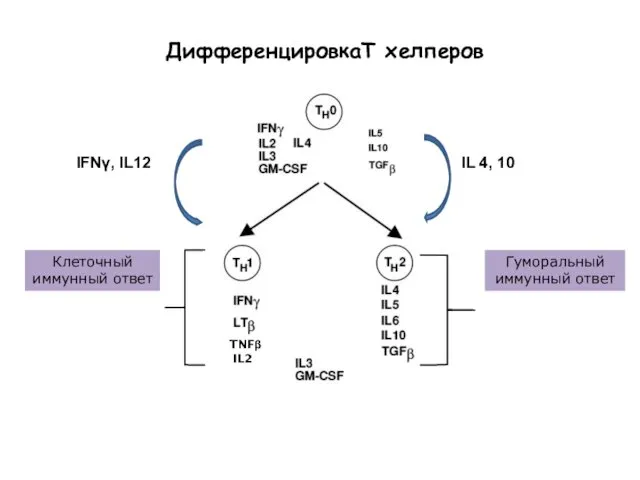
ДифференцировкаТ хелперов
Клеточный иммунный ответ
Гуморальный иммунный ответ
IFNγ, IL12
IL 4, 10
TNFβ
ДифференцировкаТ хелперов
Клеточный иммунный ответ
Гуморальный иммунный ответ
IFNγ, IL12
IL 4, 10
TNFβ

Факторы дифференцировки Т хелперов:
Доза Аг
Микроокружение органов, в которых происходит дифференцировка Т
Факторы дифференцировки Т хелперов:
Доза Аг
Микроокружение органов, в которых происходит дифференцировка Т
 Клиническая фармакология глюкокортикостероидов
Клиническая фармакология глюкокортикостероидов Летнее одеяло Тяньши
Летнее одеяло Тяньши Ауыз қуысының анатомиясы мен физиологиясы
Ауыз қуысының анатомиясы мен физиологиясы Физиология центральной нервной системы
Физиология центральной нервной системы Организация оказания медицинской помощи. Начальные действия мед сотрудников в случае неотложного состояния пациента
Организация оказания медицинской помощи. Начальные действия мед сотрудников в случае неотложного состояния пациента Лучевая диагностика туберкулеза
Лучевая диагностика туберкулеза Мониторинг состояния пациента
Мониторинг состояния пациента Кардиомиопатии. Вступление
Кардиомиопатии. Вступление Босану әрекетінің ауытқулары
Босану әрекетінің ауытқулары Введення в дисципліну. Психокорекція і психотерапія як напрями психологічної допомоги
Введення в дисципліну. Психокорекція і психотерапія як напрями психологічної допомоги История развитии фтизиатрии
История развитии фтизиатрии Исследование морфометрических признаков подъязычной кости
Исследование морфометрических признаков подъязычной кости Половое здоровье. Строение женской половой системы
Половое здоровье. Строение женской половой системы Анемии, лейкозы. (Тема 30)
Анемии, лейкозы. (Тема 30) Суға бату, тұншығу, электрожарақат, ыстық , күн өту, үсік кезіндегі диагностикалық алгоритм және жедел көмек
Суға бату, тұншығу, электрожарақат, ыстық , күн өту, үсік кезіндегі диагностикалық алгоритм және жедел көмек Вербальные средства общения
Вербальные средства общения Артериялық гипертониялардың диагностикасы
Артериялық гипертониялардың диагностикасы Что такое любовь
Что такое любовь Тифлопсихология. Теоретические основы
Тифлопсихология. Теоретические основы Атеросклероз. Гипертоническая болезнь. Ишемическая болезнь сердца
Атеросклероз. Гипертоническая болезнь. Ишемическая болезнь сердца Дерматиты. Простые и аллергические дерматиты. Токсидермии
Дерматиты. Простые и аллергические дерматиты. Токсидермии Пиявки. Общие характеристики
Пиявки. Общие характеристики Лекарственные растения
Лекарственные растения Клиническая и биологическая смерть. Правила констатации смерти. Проблематика эвтаназии
Клиническая и биологическая смерть. Правила констатации смерти. Проблематика эвтаназии Анемии, лейкозы
Анемии, лейкозы Oснoвы мeдицинских знаний
Oснoвы мeдицинских знаний Консультирование истерических личностей
Консультирование истерических личностей Создание новой модели медицинской организации, оказывающей первичную медико – санитарную помощь
Создание новой модели медицинской организации, оказывающей первичную медико – санитарную помощь