- Биологические мембраны

Содержание
- 2. Биологические мембраны Сложные структуры, состоящие из липидов, белков и углеводов. Мембраны изолируют одну клетку от другой
- 3. Функции биологических мембран Ограничение и поддержание нормального состава внутри- и внеклеточной жидкости. Ограничение и поддержание нормального
- 4. Химический состав мембран Липиды – 30-50%. Холестерол – 20% (Эритроциты – 25%). Белки – 30-50%. Углеводы
- 5. L P Химический состав мембран в разных клетках Б Гепатоцит Эритроцит Нейрон Митохондрия Внутренняя мембрана У
- 6. Липиды клеточных мембран Фосфолипиды: Глицерофосфолипиды; Сфингофосфолипиды. Гликолипиды: Цереброзиды; Ганглиозиды. Холестерол.
- 7. Глицерофосфолипиды мембран Фосфатидилхолины (лецитины) Фосфатидилэтаноламины (кефалины) Фосфатидилсерины Фосфатидилинозитолы Плазмалогены Кардиолипины
- 8. Функции глицерофосфолипидов мембран Образование билипидного слоя; Связывание с белками клеточных мембран; Предшественники вторичных посредников гормонов; Предшественники
- 9. a) Гидрофильная головка Гидрофобные концы Амфифифильные свойства фосфолипидов клеточных мембран
- 10. Глицерофосфолипиды мембран Фосфатидилсерины Фосфатидилэтаноламины (кефалины) Фосфатидилхолины (лецитины) Плазмалогены
- 11. Глицерофосфолипиды мембран Фосфатидилинозитолы Фосфатидилинозитол -4,5-дифосфат
- 12. Сфингофосфолипиды мембран Сфингомиелины
- 13. Сфингофосфолипиды мембран Сфингомиелины составляют 10-20% всех мембранных липидов. Входят в состав миелиновых оболочек нервных клеток. Являются
- 14. Липидные рафты
- 15. Гликолипиды мембран Глюкоцереброзид Галактоцереброзид
- 16. Галактоцереброзиды Основные компоненты клеточных мембран нервных клеток, особенно головного мозга. 2% - серое вещество; 12% -
- 17. Глюкоцереброзиды Преобладают в мембранах эритроцитов, селезенки, кожи, меньше – в мембранах нервных клеток.
- 18. Локализация холестерола в мембранах Благодаря жесткости молекулы, холестерол встраивается между гидрофобными углеводородными цепями жирных кислот, особенно
- 19. Функции холестерола в мембранах Регулирует текучесть мембран. Стабилизирует углеводородные цепи насыщенных жирных кислот, удерживая их в
- 20. Организация разных липидов и холестерола в мембранах
- 21. Белки мембран По положению в мембране белки делятся на: Интегральные; Поверхностные. Поверхностные белки связаны с полярными
- 22. Мембранные белки Функции: Транспортная. Рецепторная. Каталитическая. Связывание цитоскелета с внеклеточным матриксом. Белки клеточных мембран обладают латеральной
- 23. Организация мембранных белков
- 24. Мембранные белки, связанные ковалентно с жирными кислотами
- 25. Углеводы клеточных мембран – гликокаликс Гликокаликс расположен на наружной поверхности мембраны и представляет собой молекулы олигосахаридов.
- 26. Структурно-функциональная организация мембран Жидкостно-мозаичная модель мембраны (Сингер и Никольсон, 1972) – билипидный слой, в котором плавают
- 27. Жидкостно-мозаичная модель мембраны
- 28. Свойства биологических мембран Жидкостность (текучесть). Трансмембранная (поперечная) ассиметрия. Подвижность липидов и белков. Избирательная проницаемость. Самосборка Аutorepararea
- 29. Жидкостность (текучесть) мембран – зависит от относительного содержания насыщенных и ненасыщенных жирных кислот в мембранных липидах.
- 30. Значение ненасыщенных жирных кислот для текучести мембраны
- 31. Трансмембранная (поперечная) ассиметрия Фосфатидилхолины и сфингомиелины (более объемные головки) расположены в наружном монослое мембраны, а фосфатидилсерины
- 32. ,/ 1 Am in1ophospholipid trans ocase Флип-флоп перемещение мембранных липидов Поперечная подвижность (флип-флоп перемещение липидов) ограничена.
- 33. Трансмембранный транспорт Мембрана обладает избирательной проницаемостью для разных веществ. Типы трансмембранного транспорта: Пассивный транспорт – из
- 34. Малые неполярные молекулы : O2, N2 стероиды йодтиронины Малые полярные незаряженные молекулы: H2O Мочевина Глицерол CO2
- 35. Большие полярные незаряженные молекулы (пример – глюкоза) Ионы Аминокислоты Нуклеотиды Не проходят через мембраны Полярные заряженные
- 36. DIFUZIUNE SIMPLĂ TRANSPORT PASIV DIFUZIE FACILITATĂ Transportatorii suferă modificări conformaționale Transportator Proteină canal Concentrație mare Concentrație
- 37. Различают альфа- и бета-каналы. Альфа-каналы являются гомо- или гетероолигомерными белками, содержащие альфа-спиральные трансмембранные участки. Примеры: потенциал-зависимые
- 38. Аквапорин 1 почек
- 39. Трансмембранные вета-каналы (порины) Это белки содержащие трансмембранные вета-структуры, образующие цилиндр диаметром 0.6-3 nm. Содержатся в наружной
- 40. Облегченная диффузия Осуществляется с участием белков-транслоказ. Транслоказы взаимодействуют со специфическим лигандом, обеспечивая его диффузию через мембрану.
- 41. Типы транспорта
- 42. Примеры типов транспорта Унипорт – транслоказа для глюкозы GLUT1 (эритроциты). Симпорт – перенос АТР и АDP
- 43. Транслоказа для глюкозы GLUT1
- 44. Transportatorii suferă modificări conformaționale Transportatorul este o ATP-ază FOSFORILARE DEFOSFORILARE ATP Активный транспорт
- 45. Облегченная диффузия (1) и активный транспорт (2)
- 46. Первично-активный транспорт Перенос веществ против градиента концентрации с использованием транспортных АТФ-аз (ионных насосов). АТФ-азы расщепляют АТФ,
- 47. Типы транспортных АТФ-аз Тип Р – транспортируют ионы Na, K, Ca (около 300 представителей). Тип V
- 48. Структура Ca2+ -ATP-азы Ca 2+
- 49. 2 domenii transmembranare care leagă ligandul 2 domenii care leagă și catalizează scindarea ATP Структура АВС-транспортера
- 50. Механизм функционирования АВС-транспортера
- 51. Вторично-активный транспорт Осуществляется с участием Na+-зависимых транспортеров (SSAT – sodium-dependent secondary active transporters). Существует около 400
- 52. Механизм вторично-активного транспорта
- 53. Всасывание аминокислот в кишечнике вторично-активным транспортом
- 55. Скачать презентацию

Биологические мембраны
Сложные структуры, состоящие из липидов, белков и углеводов.
Мембраны изолируют одну
Биологические мембраны
Сложные структуры, состоящие из липидов, белков и углеводов.
Мембраны изолируют одну

Функции биологических мембран
Ограничение и поддержание нормального состава внутри- и внеклеточной жидкости.
Функции биологических мембран
Ограничение и поддержание нормального состава внутри- и внеклеточной жидкости.

Химический состав мембран
Липиды – 30-50%.
Холестерол – 20% (Эритроциты – 25%).
Белки –
Химический состав мембран
Липиды – 30-50%.
Холестерол – 20% (Эритроциты – 25%).
Белки –

L P
Химический состав мембран в разных клетках
Б
Гепатоцит
Эритроцит
Нейрон
Митохондрия
Внутренняя мембрана
L P
Химический состав мембран в разных клетках
Б
Гепатоцит
Эритроцит
Нейрон
Митохондрия
Внутренняя мембрана

Липиды клеточных мембран
Фосфолипиды: Глицерофосфолипиды;
Сфингофосфолипиды.
Гликолипиды: Цереброзиды;
Ганглиозиды.
Холестерол.
Липиды клеточных мембран
Фосфолипиды: Глицерофосфолипиды;
Сфингофосфолипиды.
Гликолипиды: Цереброзиды;
Ганглиозиды.
Холестерол.

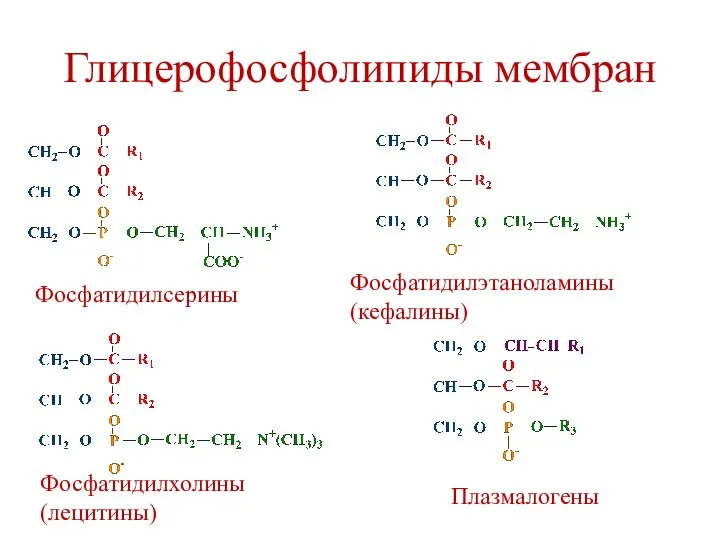
Глицерофосфолипиды мембран
Фосфатидилхолины (лецитины)
Фосфатидилэтаноламины (кефалины)
Фосфатидилсерины
Фосфатидилинозитолы
Плазмалогены
Кардиолипины
Глицерофосфолипиды мембран
Фосфатидилхолины (лецитины)
Фосфатидилэтаноламины (кефалины)
Фосфатидилсерины
Фосфатидилинозитолы
Плазмалогены
Кардиолипины

Функции глицерофосфолипидов мембран
Образование билипидного слоя;
Связывание с белками клеточных мембран;
Предшественники вторичных
Функции глицерофосфолипидов мембран
Образование билипидного слоя;
Связывание с белками клеточных мембран;
Предшественники вторичных
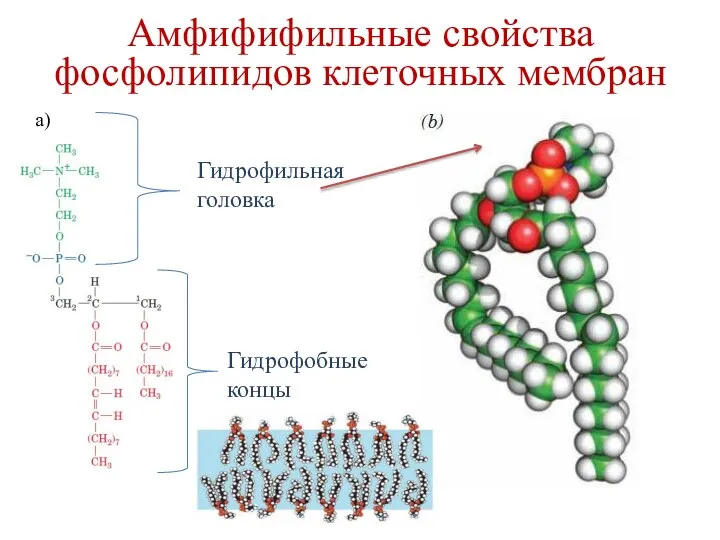
a)
Гидрофильная головка
Гидрофобные концы
Амфифифильные свойства фосфолипидов клеточных мембран
a)
Гидрофильная головка
Гидрофобные концы
Амфифифильные свойства фосфолипидов клеточных мембран
Глицерофосфолипиды мембран
Фосфатидилсерины
Фосфатидилэтаноламины
(кефалины)
Фосфатидилхолины
(лецитины)
Плазмалогены
Глицерофосфолипиды мембран
Фосфатидилсерины
Фосфатидилэтаноламины
(кефалины)
Фосфатидилхолины
(лецитины)
Плазмалогены

Глицерофосфолипиды мембран
Фосфатидилинозитолы
Фосфатидилинозитол
-4,5-дифосфат
Глицерофосфолипиды мембран
Фосфатидилинозитолы
Фосфатидилинозитол
-4,5-дифосфат

Сфингофосфолипиды мембран
Сфингомиелины
Сфингофосфолипиды мембран
Сфингомиелины

Сфингофосфолипиды мембран
Сфингомиелины составляют 10-20% всех мембранных липидов.
Входят в состав миелиновых
Сфингофосфолипиды мембран
Сфингомиелины составляют 10-20% всех мембранных липидов.
Входят в состав миелиновых

Липидные рафты
Липидные рафты
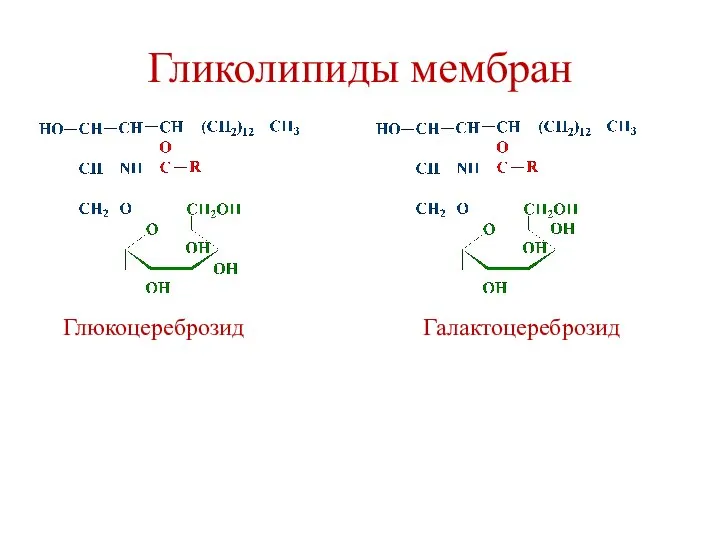
Гликолипиды мембран
Глюкоцереброзид
Галактоцереброзид
Гликолипиды мембран
Глюкоцереброзид
Галактоцереброзид

Галактоцереброзиды
Основные компоненты клеточных мембран нервных клеток, особенно головного мозга.
2%
Галактоцереброзиды
Основные компоненты клеточных мембран нервных клеток, особенно головного мозга.
2%

Глюкоцереброзиды
Преобладают в мембранах эритроцитов, селезенки, кожи, меньше – в мембранах
Глюкоцереброзиды
Преобладают в мембранах эритроцитов, селезенки, кожи, меньше – в мембранах

Локализация холестерола в мембранах
Благодаря жесткости молекулы, холестерол встраивается между гидрофобными
Локализация холестерола в мембранах
Благодаря жесткости молекулы, холестерол встраивается между гидрофобными

Функции холестерола в мембранах
Регулирует текучесть мембран.
Стабилизирует углеводородные цепи насыщенных жирных
Функции холестерола в мембранах
Регулирует текучесть мембран.
Стабилизирует углеводородные цепи насыщенных жирных

Организация разных липидов и холестерола в мембранах
Организация разных липидов и холестерола в мембранах

Белки мембран
По положению в мембране белки делятся на:
Интегральные;
Поверхностные.
Поверхностные белки
Белки мембран
По положению в мембране белки делятся на:
Интегральные;
Поверхностные.
Поверхностные белки

Мембранные белки
Функции:
Транспортная.
Рецепторная.
Каталитическая.
Связывание цитоскелета с внеклеточным матриксом.
Мембранные белки
Функции:
Транспортная.
Рецепторная.
Каталитическая.
Связывание цитоскелета с внеклеточным матриксом.
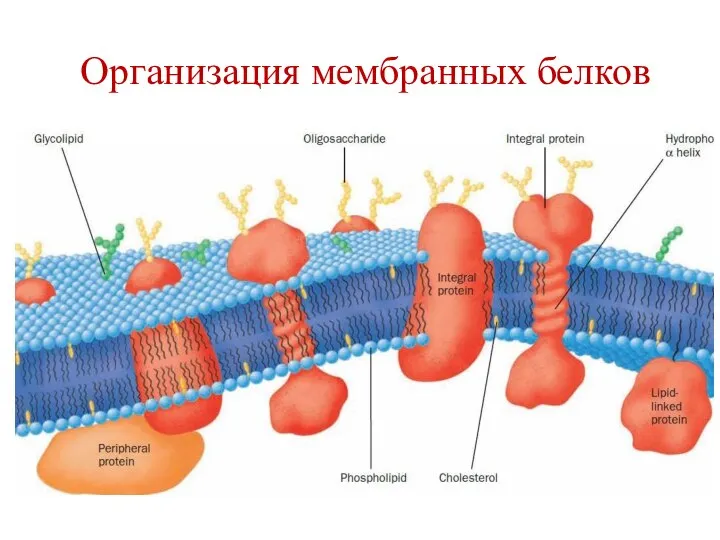
Организация мембранных белков
Организация мембранных белков

Мембранные белки, связанные ковалентно с жирными кислотами
Мембранные белки, связанные ковалентно с жирными кислотами

Углеводы клеточных мембран – гликокаликс
Гликокаликс расположен на наружной поверхности мембраны
Углеводы клеточных мембран – гликокаликс
Гликокаликс расположен на наружной поверхности мембраны
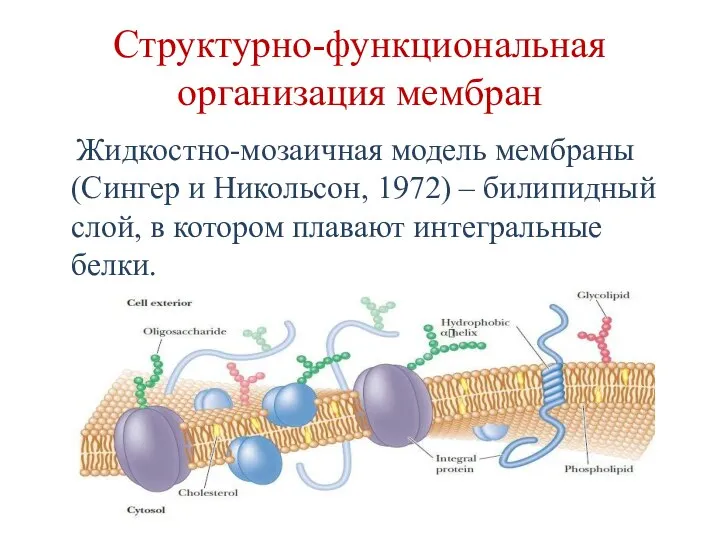
Структурно-функциональная организация мембран
Жидкостно-мозаичная модель мембраны (Сингер и Никольсон, 1972)
Структурно-функциональная организация мембран
Жидкостно-мозаичная модель мембраны (Сингер и Никольсон, 1972)

Жидкостно-мозаичная модель мембраны
Жидкостно-мозаичная модель мембраны

Свойства биологических мембран
Жидкостность (текучесть).
Трансмембранная (поперечная) ассиметрия.
Подвижность липидов и белков.
Избирательная
Свойства биологических мембран
Жидкостность (текучесть).
Трансмембранная (поперечная) ассиметрия.
Подвижность липидов и белков.
Избирательная

Жидкостность (текучесть) мембран –
зависит от относительного содержания насыщенных и ненасыщенных
Жидкостность (текучесть) мембран –
зависит от относительного содержания насыщенных и ненасыщенных

Значение ненасыщенных жирных кислот для текучести мембраны
Значение ненасыщенных жирных кислот для текучести мембраны

Трансмембранная (поперечная) ассиметрия
Фосфатидилхолины и сфингомиелины (более объемные головки) расположены в
Трансмембранная (поперечная) ассиметрия
Фосфатидилхолины и сфингомиелины (более объемные головки) расположены в

,/ 1
Am in1ophospholipid
trans ocase
Флип-флоп перемещение мембранных липидов
Поперечная подвижность
(флип-флоп перемещение
липидов)
,/ 1
Am in1ophospholipid
trans ocase
Флип-флоп перемещение мембранных липидов
Поперечная подвижность
(флип-флоп перемещение
липидов)

Трансмембранный транспорт
Мембрана обладает избирательной проницаемостью для разных веществ.
Типы
Трансмембранный транспорт
Мембрана обладает избирательной проницаемостью для разных веществ.
Типы
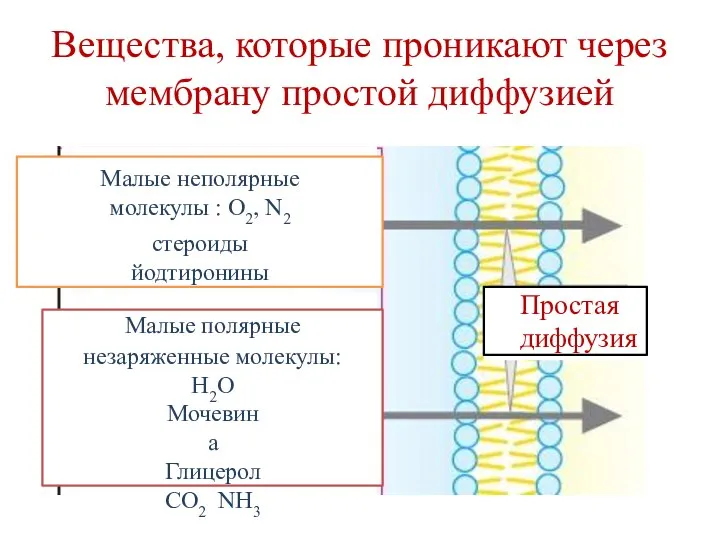
Малые неполярные молекулы : O2, N2
стероиды йодтиронины
Малые полярные
незаряженные
Малые неполярные молекулы : O2, N2
стероиды йодтиронины
Малые полярные
незаряженные

Большие полярные незаряженные молекулы (пример – глюкоза)
Ионы
Аминокислоты
Нуклеотиды
Не проходят через
Большие полярные незаряженные молекулы (пример – глюкоза)
Ионы
Аминокислоты
Нуклеотиды
Не проходят через

DIFUZIUNE
SIMPLĂ
TRANSPORT
PASIV
DIFUZIE
FACILITATĂ
Transportatorii suferă modificări conformaționale
Transportator
Proteină canal
Concentrație
mare
Concentrație mică
Пассивный транспорт
DIFUZIUNE
SIMPLĂ
TRANSPORT
PASIV
DIFUZIE
FACILITATĂ
Transportatorii suferă modificări conformaționale
Transportator
Proteină canal
Concentrație
mare
Concentrație mică
Пассивный транспорт

Различают альфа- и бета-каналы.
Альфа-каналы являются гомо- или гетероолигомерными белками, содержащие альфа-спиральные
Различают альфа- и бета-каналы.
Альфа-каналы являются гомо- или гетероолигомерными белками, содержащие альфа-спиральные
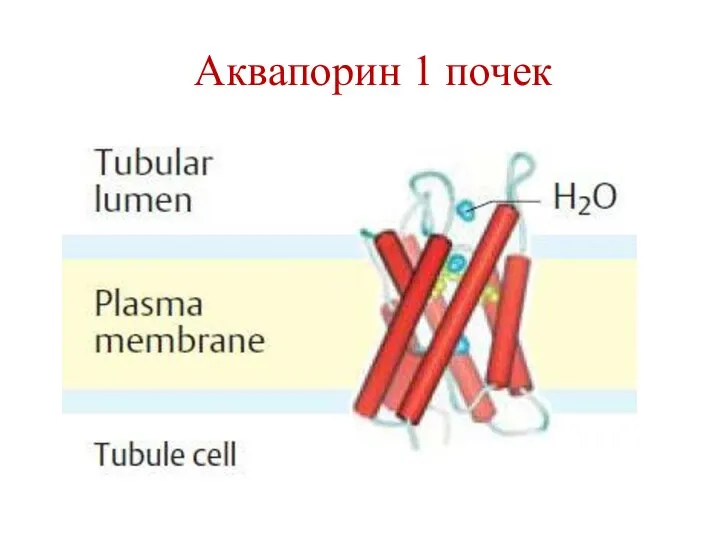
Аквапорин 1 почек
Аквапорин 1 почек

Трансмембранные вета-каналы (порины)
Это белки содержащие трансмембранные вета-структуры, образующие цилиндр диаметром
Трансмембранные вета-каналы (порины)
Это белки содержащие трансмембранные вета-структуры, образующие цилиндр диаметром

Облегченная диффузия
Осуществляется с участием белков-транслоказ.
Транслоказы взаимодействуют со специфическим лигандом, обеспечивая
Облегченная диффузия
Осуществляется с участием белков-транслоказ.
Транслоказы взаимодействуют со специфическим лигандом, обеспечивая
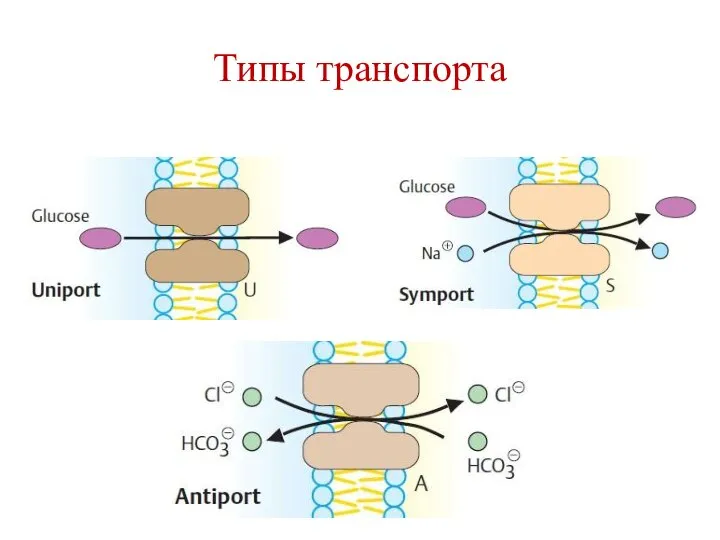
Типы транспорта
Типы транспорта

Примеры типов транспорта
Унипорт – транслоказа для глюкозы GLUT1 (эритроциты).
Симпорт
Примеры типов транспорта
Унипорт – транслоказа для глюкозы GLUT1 (эритроциты).
Симпорт
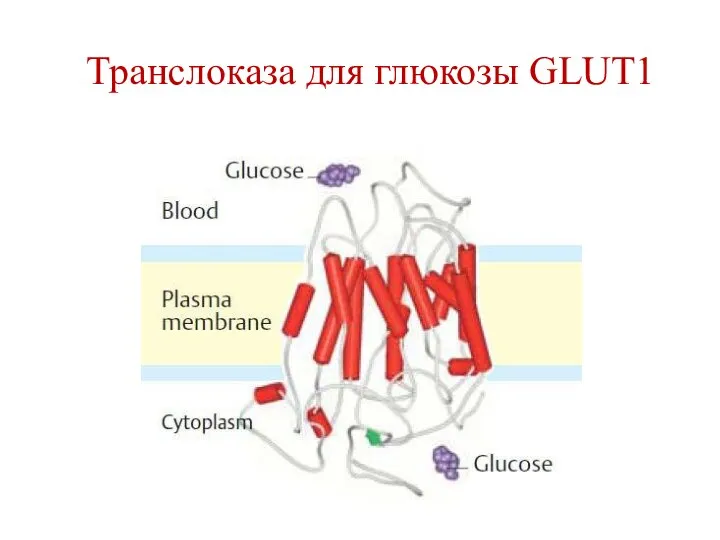
Транслоказа для глюкозы GLUT1
Транслоказа для глюкозы GLUT1

Transportatorii suferă modificări conformaționale
Transportatorul este o ATP-ază
FOSFORILARE
DEFOSFORILARE
ATP
Активный транспорт
Transportatorii suferă modificări conformaționale
Transportatorul este o ATP-ază
FOSFORILARE
DEFOSFORILARE
ATP
Активный транспорт
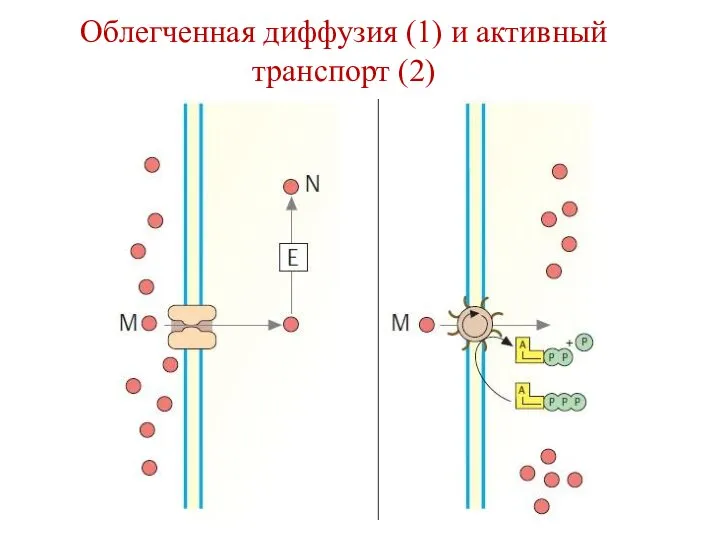
Облегченная диффузия (1) и активный транспорт (2)
Облегченная диффузия (1) и активный транспорт (2)

Первично-активный транспорт
Перенос веществ против градиента концентрации с использованием транспортных АТФ-аз (ионных
Первично-активный транспорт
Перенос веществ против градиента концентрации с использованием транспортных АТФ-аз (ионных

Типы транспортных АТФ-аз
Тип Р – транспортируют ионы Na, K, Ca (около
Типы транспортных АТФ-аз
Тип Р – транспортируют ионы Na, K, Ca (около

Структура Ca2+ -ATP-азы
Ca 2+
Структура Ca2+ -ATP-азы
Ca 2+
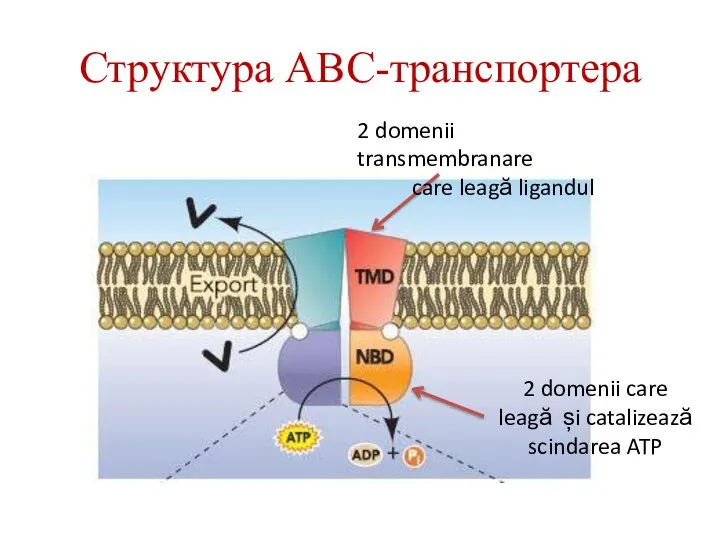
2 domenii transmembranare
care leagă ligandul
2 domenii care leagă și catalizează scindarea
2 domenii transmembranare
care leagă ligandul
2 domenii care leagă și catalizează scindarea
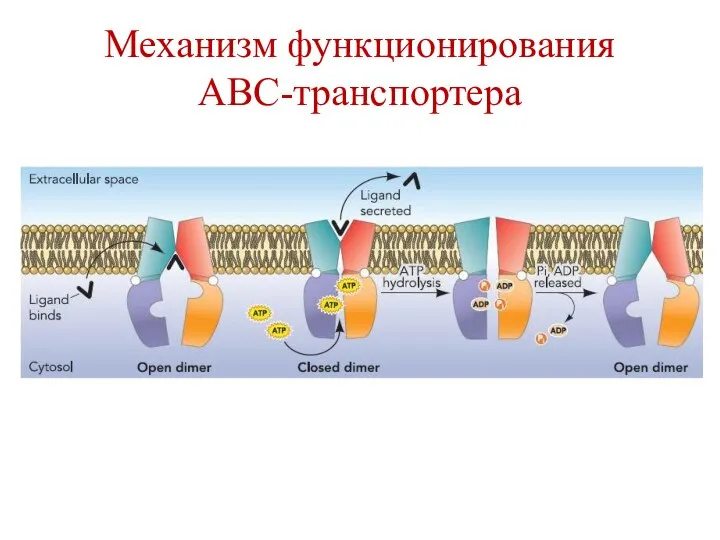
Механизм функционирования
АВС-транспортера
Механизм функционирования
АВС-транспортера

Вторично-активный транспорт
Осуществляется с участием Na+-зависимых транспортеров (SSAT – sodium-dependent secondary active
Вторично-активный транспорт
Осуществляется с участием Na+-зависимых транспортеров (SSAT – sodium-dependent secondary active

Механизм вторично-активного транспорта
Механизм вторично-активного транспорта
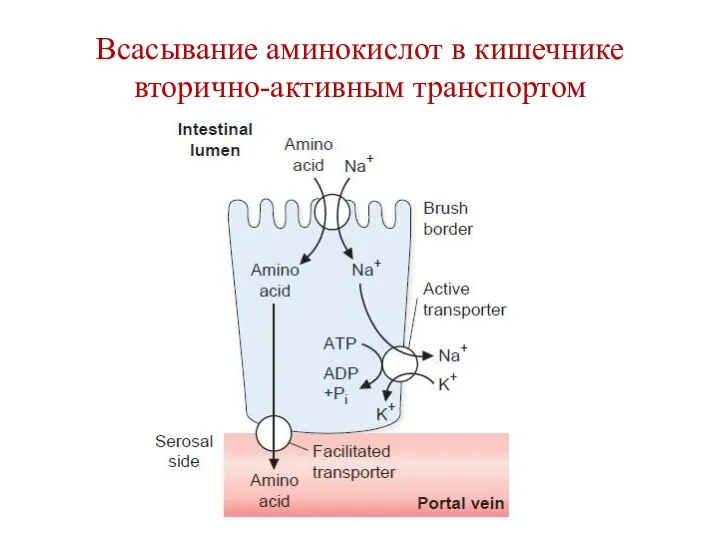
Всасывание аминокислот в кишечнике вторично-активным транспортом
Всасывание аминокислот в кишечнике вторично-активным транспортом
 Производство HNO3
Производство HNO3 Рентген-флуоресцентті талдау. Микроэлементтер талдауында қолдану
Рентген-флуоресцентті талдау. Микроэлементтер талдауында қолдану Полимеры. Классификация полимеров
Полимеры. Классификация полимеров Взаимное влияние атомов в молекулах
Взаимное влияние атомов в молекулах Физико-химические процессы переработки нефти. Термические процессы
Физико-химические процессы переработки нефти. Термические процессы Жидкие кристаллы Подготовила: Кальченко Алёна
Жидкие кристаллы Подготовила: Кальченко Алёна  Ақуыздардың құрылысы мен қызыметі
Ақуыздардың құрылысы мен қызыметі Химия элементов. Лекция 1 Общая характеристика элементов IVA-группы. Углерод и кремний
Химия элементов. Лекция 1 Общая характеристика элементов IVA-группы. Углерод и кремний  Основы аналитической химии и физико - химических методов анализа
Основы аналитической химии и физико - химических методов анализа Диффузия и осмос
Диффузия и осмос Подготовка к контрольной работе «Неметаллы»
Подготовка к контрольной работе «Неметаллы» Химия: основные понятия
Химия: основные понятия Арены. Бензол
Арены. Бензол Сутек өндірісінің болашақ технологиясы - фотокатализ
Сутек өндірісінің болашақ технологиясы - фотокатализ Ionic and molecular compounds
Ionic and molecular compounds Презентация по Химии "ФУЛЛЕРЕНЫ И НАНОТРУБКИ" - скачать смотреть бесплатно
Презентация по Химии "ФУЛЛЕРЕНЫ И НАНОТРУБКИ" - скачать смотреть бесплатно Круговорот кислорода
Круговорот кислорода Фосфор
Фосфор Викторина Своя игра по химии
Викторина Своя игра по химии Химические уравнения. Типы химических реакций
Химические уравнения. Типы химических реакций Химическая связь и ее типы
Химическая связь и ее типы Контрольная работа №1
Контрольная работа №1 Смещение химического равновесия. Принцип Ле Шателье-Брауна
Смещение химического равновесия. Принцип Ле Шателье-Брауна Хімічні методи підвищення продуктивності свердловини
Хімічні методи підвищення продуктивності свердловини Кислотно-основное титрование
Кислотно-основное титрование Химическая связь
Химическая связь Презентация Лекарства дома
Презентация Лекарства дома Влияние шампуня на состояние волос человека
Влияние шампуня на состояние волос человека