- Признаки рака

Содержание
- 2. Нестабильность генома Поддержание пролиферативного сигналинга Сопротивление клеточной гибели Неограниченное деление Индуцирование ангиогенеза Активация инвазии и метастазирования
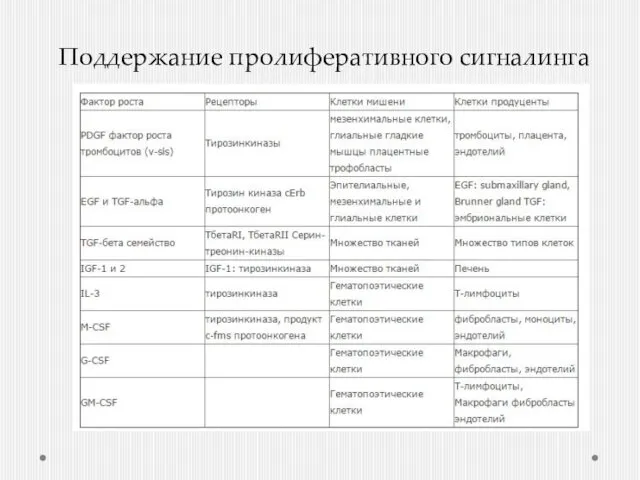
- 3. Поддержание пролиферативного сигналинга
- 4. Фактор роста Активация РТК Синтез белков S-фаза Митоз Каскад фосфорилирования Факторы транскрипции Перепрограммирование генома Синтез мРНК
- 5. Как поддержать пролиферативный сигнал? Производство собственных факторов роста, на которые сами же клетки-производители и будут реагировать
- 6. Гиперэкспрессия рецепторов EGFR – 60% НМРЛ, 50% - глиома Erb2 – 20-30% РМЖ Амплификация гена Повышение
- 7. К чему приводит мутация B-Raf белка?
- 8. Активация PI3K (фосфоинозитол-3-киназы)
- 9. Нарушение механизма отрицательной обратной связи ГТФаза
- 10. Существуют ли меры борьбы клетки с чрезмерным пролиферативным сигналингом?
- 11. Нестабильность генома Поддержание пролиферативного сигналинга Сопротивление клеточной гибели Неограниченное деление Индуцирование ангиогенеза Активация инвазии и метастазирования
- 12. Онкосупрессоры RB-белок (белок ретинобластомы) «решает, пройти ли клетке через следующий цикл деления» мутация Неконтролируемая клеточная пролиферация
- 13. Контактное торможение Нормальная популяция клеток in-vitro при действии агрессивного фактора образуют слившийся монослой и приостанавливают пролиферацию.
- 14. Механизмы контактного торможения Cвязывание молекул адгезии (Е-кадгерина) с РТК (EGFR) * «занимает» РТК => ограничение ответа
- 15. Нестабильность генома Поддержание пролиферативного сигналинга Сопротивление клеточной гибели Неограниченное деление Индуцирование ангиогенеза Активация инвазии и метастазирования
- 16. Апоптоз Внешние сигналы Внутренние сигналы Активация «молчащих» протеаз (каспаз 8,9) Протеолиз Фагоцитоз Вах и Вас белки
- 17. Пути избегания апоптоза Внешние сигналы Внутренние сигналы Активация «молчащих» протеаз (каспаз 8,9) Протеолиз Фагоцитоз Вах и
- 18. Роль некроза клеток в стимуляции онкогенеза Клетки, вступившие в некроз привлекают в очаг элементы иммунной системы
- 19. Нестабильность генома Поддержание пролиферативного сигналинга Сопротивление клеточной гибели Неограниченное деление Индуцирование ангиогенеза Активация инвазии и метастазирования
- 20. «Клеточное бессмертие» эффект Хейфлика (теломерная теория) теломераза (экспрессируется в 90% опухолевых клетках) Удлинняет теломерную ДНК Стимулирует
- 21. Нестабильность генома Поддержание пролиферативного сигналинга Сопротивление клеточной гибели Неограниченное деление Индуцирование ангиогенеза Активация инвазии и метастазирования
- 22. Проангиогенный эффект VEGF (vascular endothelial growth factor) FGF (фактор роста фибробластов) Антиангиогенный эффект TSP-1 (тромбоспондин) Коллаген
- 23. Роль клеток иммунной системы в ангиогенезе Макрофаги, нейтрофилы, тучные клетки и миелоидные предшественники, инфильтрирующие предраковые очаги,
- 24. Нестабильность генома Поддержание пролиферативного сигналинга Сопротивление клеточной гибели Неограниченное деление Индуцирование ангиогенеза Активация инвазии и метастазирования
- 25. Механизмы инвазии Нарушение межклеточного взаимодействия путем мутации в молекулах адгезии (в основном в Е-кадгерине) Виды инвазии:
- 26. Эпителиально-мезенхимальный переход Подавление экспрессии гена Е-кадгерина (E-cadherin (CDH1)) Увеличение экспрессии генов ответственных за мезенхимальный фенотип эпителиоцитов,
- 27. - Транскрипционные факторы, участвующие в ЭМП способны контролировать все этапы, исключая заключительный. - Опухолевые клетки выделяют
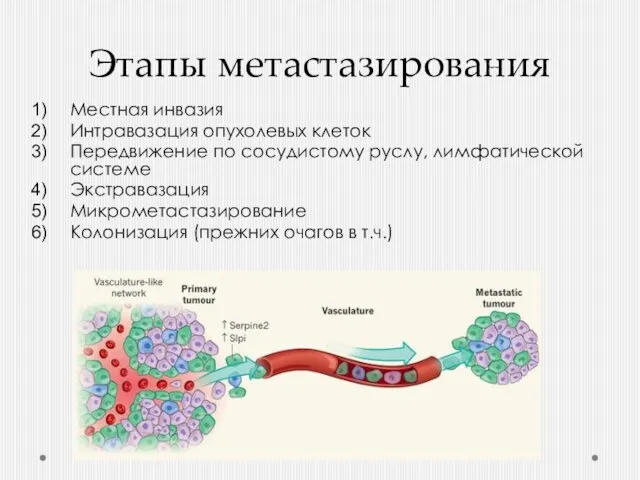
- 28. Этапы метастазирования Местная инвазия Интравазация опухолевых клеток Передвижение по сосудистому руслу, лимфатической системе Экстравазация Микрометастазирование Колонизация
- 29. Мигрирующие клетки могут циркулировать в русле годами и, попав на место первичной опухоли, сформировать фенотипически совершенно
- 30. Нестабильность генома Поддержание пролиферативного сигналинга Сопротивление клеточной гибели Неограниченное деление Индуцирование ангиогенеза Активация инвазии и метастазирования
- 31. Репрограммирование энергетического потенциала Активация онкогенов и мутация опухолевых супрессоров Аэробный гликолиз Увеличение активности переносчика глюкозы (GLUT-1)
- 32. В чем смысл 18-кратного снижения эффективности образования АТФ? Использование промежуточных метаболитов гликолиза в различные биосинтетические пути,
- 35. Скачать презентацию

Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
Поддержание пролиферативного сигналинга
Поддержание пролиферативного сигналинга
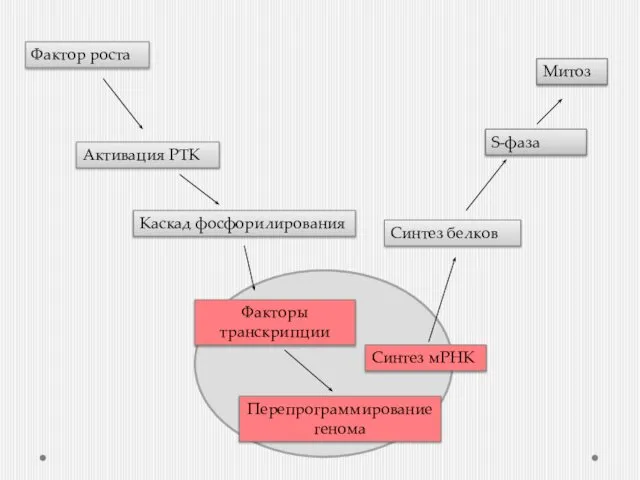
Фактор роста
Активация РТК
Синтез белков
S-фаза
Митоз
Каскад фосфорилирования
Факторы транскрипции
Перепрограммирование генома
Синтез мРНК
Фактор роста
Активация РТК
Синтез белков
S-фаза
Митоз
Каскад фосфорилирования
Факторы транскрипции
Перепрограммирование генома
Синтез мРНК

Как поддержать пролиферативный сигнал?
Производство собственных факторов роста, на которые сами же
Как поддержать пролиферативный сигнал?
Производство собственных факторов роста, на которые сами же

Гиперэкспрессия рецепторов
EGFR – 60% НМРЛ, 50% - глиома
Erb2 – 20-30%
Гиперэкспрессия рецепторов
EGFR – 60% НМРЛ, 50% - глиома
Erb2 – 20-30%
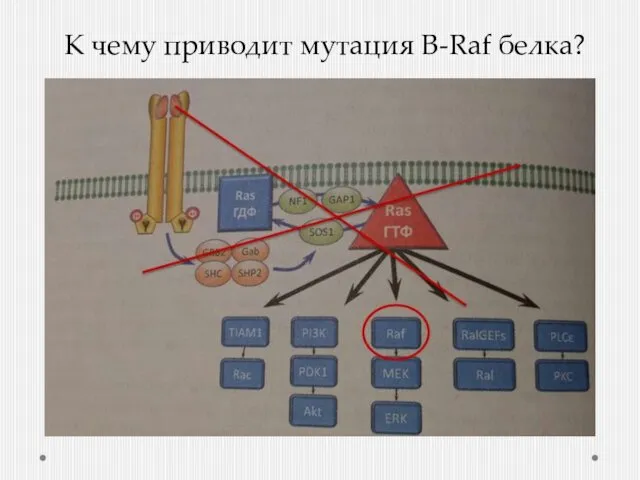
К чему приводит мутация B-Raf белка?
К чему приводит мутация B-Raf белка?

Активация PI3K
(фосфоинозитол-3-киназы)
Активация PI3K
(фосфоинозитол-3-киназы)

Нарушение механизма отрицательной обратной связи
ГТФаза
Нарушение механизма отрицательной обратной связи
ГТФаза
Существуют ли меры борьбы клетки с чрезмерным пролиферативным сигналингом?
Существуют ли меры борьбы клетки с чрезмерным пролиферативным сигналингом?

Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение

Онкосупрессоры
RB-белок
(белок ретинобластомы) «решает, пройти ли клетке через следующий цикл деления»
Онкосупрессоры
RB-белок
(белок ретинобластомы) «решает, пройти ли клетке через следующий цикл деления»

Контактное торможение
Нормальная популяция клеток in-vitro при действии агрессивного фактора образуют слившийся
Контактное торможение
Нормальная популяция клеток in-vitro при действии агрессивного фактора образуют слившийся

Механизмы контактного торможения
Cвязывание молекул адгезии (Е-кадгерина) с РТК (EGFR)
* «занимает» РТК
Механизмы контактного торможения
Cвязывание молекул адгезии (Е-кадгерина) с РТК (EGFR)
* «занимает» РТК

Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
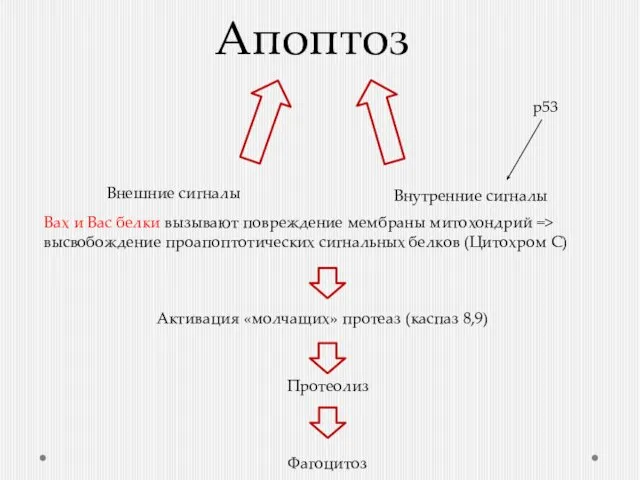
Апоптоз
Внешние сигналы
Внутренние сигналы
Активация «молчащих» протеаз (каспаз 8,9)
Протеолиз
Фагоцитоз
Вах и
Апоптоз
Внешние сигналы
Внутренние сигналы
Активация «молчащих» протеаз (каспаз 8,9)
Протеолиз
Фагоцитоз
Вах и
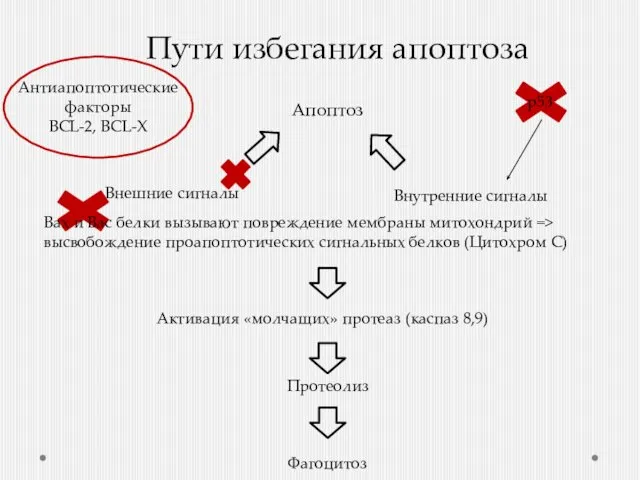
Пути избегания апоптоза
Внешние сигналы
Внутренние сигналы
Активация «молчащих» протеаз (каспаз 8,9)
Протеолиз
Пути избегания апоптоза
Внешние сигналы
Внутренние сигналы
Активация «молчащих» протеаз (каспаз 8,9)
Протеолиз

Роль некроза клеток в стимуляции онкогенеза
Клетки, вступившие в некроз привлекают в
Роль некроза клеток в стимуляции онкогенеза
Клетки, вступившие в некроз привлекают в

Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
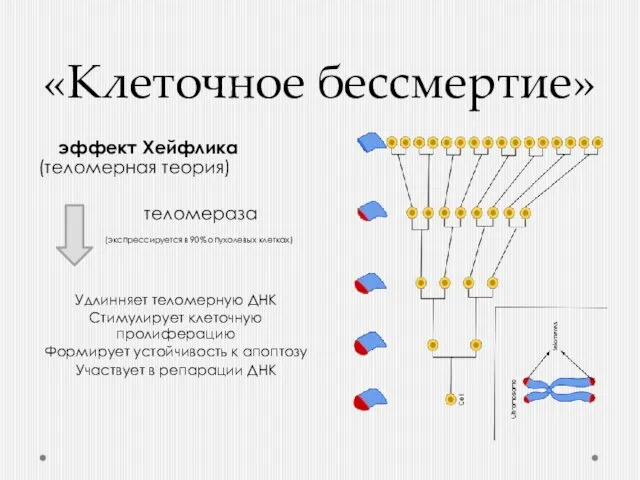
«Клеточное бессмертие»
эффект Хейфлика (теломерная теория)
теломераза
(экспрессируется в 90% опухолевых
«Клеточное бессмертие»
эффект Хейфлика (теломерная теория)
теломераза
(экспрессируется в 90% опухолевых

Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение

Проангиогенный эффект
VEGF (vascular endothelial growth factor)
FGF (фактор роста фибробластов)
Антиангиогенный
Проангиогенный эффект
VEGF (vascular endothelial growth factor)
FGF (фактор роста фибробластов)
Антиангиогенный

Роль клеток иммунной системы в ангиогенезе
Макрофаги, нейтрофилы, тучные клетки и
Роль клеток иммунной системы в ангиогенезе
Макрофаги, нейтрофилы, тучные клетки и

Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение

Механизмы инвазии
Нарушение межклеточного взаимодействия путем мутации в молекулах адгезии (в основном
Механизмы инвазии
Нарушение межклеточного взаимодействия путем мутации в молекулах адгезии (в основном

Эпителиально-мезенхимальный переход
Подавление экспрессии гена Е-кадгерина (E-cadherin (CDH1))
Увеличение экспрессии генов ответственных
Эпителиально-мезенхимальный переход
Подавление экспрессии гена Е-кадгерина (E-cadherin (CDH1))
Увеличение экспрессии генов ответственных
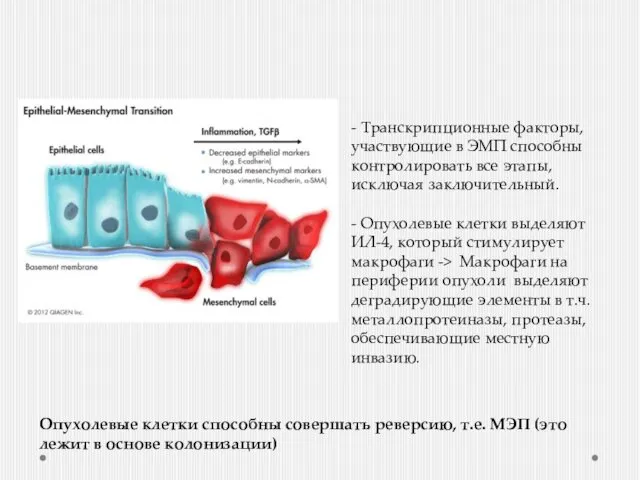
- Транскрипционные факторы, участвующие в ЭМП способны контролировать все этапы, исключая
- Транскрипционные факторы, участвующие в ЭМП способны контролировать все этапы, исключая
Этапы метастазирования
Местная инвазия
Интравазация опухолевых клеток
Передвижение по сосудистому руслу, лимфатической
Этапы метастазирования
Местная инвазия
Интравазация опухолевых клеток
Передвижение по сосудистому руслу, лимфатической

Мигрирующие клетки могут циркулировать в русле годами и, попав на место
Мигрирующие клетки могут циркулировать в русле годами и, попав на место

Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
Нестабильность генома
Поддержание пролиферативного сигналинга
Сопротивление клеточной гибели
Неограниченное деление
Индуцирование ангиогенеза
Активация инвазии и метастазирования
Избегаение
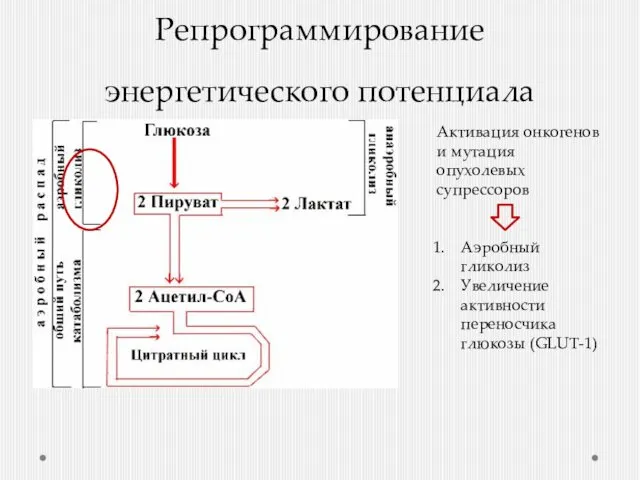
Репрограммирование энергетического потенциала
Активация онкогенов и мутация опухолевых супрессоров
Аэробный гликолиз
Увеличение активности
Репрограммирование энергетического потенциала
Активация онкогенов и мутация опухолевых супрессоров
Аэробный гликолиз
Увеличение активности

В чем смысл 18-кратного снижения эффективности образования АТФ?
Использование промежуточных метаболитов гликолиза
В чем смысл 18-кратного снижения эффективности образования АТФ?
Использование промежуточных метаболитов гликолиза

 Методы исследования генетики человека
Методы исследования генетики человека Зағиптарға арналған шрифт
Зағиптарға арналған шрифт Сөйлеу мүшелерінің анатомиясы
Сөйлеу мүшелерінің анатомиясы Калликреин-кининовая система. Прошлое и настоящее
Калликреин-кининовая система. Прошлое и настоящее Индивидная дифференциация темперамент
Индивидная дифференциация темперамент Консультирование асоциальных личностей
Консультирование асоциальных личностей возрастные особенности внутренней картины болезни
возрастные особенности внутренней картины болезни Учение о динамической локализации высших психических функций
Учение о динамической локализации высших психических функций Форменные элементы крови
Форменные элементы крови Оси и плоскости человеческого тела. Основные анатомические термины
Оси и плоскости человеческого тела. Основные анатомические термины Неврологические аспекты адаптации детей в школе, пути профилактики
Неврологические аспекты адаптации детей в школе, пути профилактики Алергічні хвороби: етіологія, унопатогенез, клініка, діагностика, лікування
Алергічні хвороби: етіологія, унопатогенез, клініка, діагностика, лікування История развития эпидемиологической службы в Рязанской области
История развития эпидемиологической службы в Рязанской области Мотивация. Потребность в широком смысле
Мотивация. Потребность в широком смысле Анатомия слезной железы. Слезная пленка. Синдром сухого глаза
Анатомия слезной железы. Слезная пленка. Синдром сухого глаза Аритмии сердца
Аритмии сердца Паразитарные заболевания глаз
Паразитарные заболевания глаз СПРУ - тяжелое хроническое заболевание
СПРУ - тяжелое хроническое заболевание Подходы к семейному консультированию. Особенности работы
Подходы к семейному консультированию. Особенности работы Бруцеллёз
Бруцеллёз Улучшение результатов хирургического лечения опухолей ободочной кишки с использованием фармакологического прекондиционирования
Улучшение результатов хирургического лечения опухолей ободочной кишки с использованием фармакологического прекондиционирования Депрессивные неврозы
Депрессивные неврозы Микробиологическая диагностика бактериемий, сепсиса
Микробиологическая диагностика бактериемий, сепсиса Головные и лицевые боли. Анатомия лицевого и тройничного нервов. Невралгия тройничного нерва, диагностика и лечение
Головные и лицевые боли. Анатомия лицевого и тройничного нервов. Невралгия тройничного нерва, диагностика и лечение Наследственные заболевания сетчатки у детей
Наследственные заболевания сетчатки у детей Подросток. Родительское собрание для 7 класса
Подросток. Родительское собрание для 7 класса Синдром нарушения ритма сердца
Синдром нарушения ритма сердца Приемы расположения к себе
Приемы расположения к себе