- Радиационное поражение живой клетки

Содержание
- 2. Стадии лучевого поражения клетки Физическая (неспецифическая) (10-16-10-14 с) – поглощение, перераспределение и деградация поглощенной энергии: возникновение
- 3. Стадии лучевого поражения клетки Физико-химическая (10-13-10-10 с) – реакции заряженных и возбужденных частиц, миграция энергии внутри
- 4. Стадии лучевого поражения клетки Химическая (10-7-10-6 с) – к этому времени уже произошли стойкие изменения –
- 5. Стадии лучевого поражения клетки Биологическая – функциональные нарушения, формирование ответных реакций клетки: Перенос против градиента (активный)
- 6. Биологическая стадия (продолжение) Структурные повреждения нуклеиновых кислот приводят к нарушению процессов репликации, транскрипции, трансляции генетической информации,
- 7. Стадии действия излучений – обобщенная схема (Цыб и др., с.67) физич Физ-хим химич биолог
- 8. Изменение радиочувствительности на разных стациях клеточного цикла Митотичеcкии цикл клетки М – митоз; G0 – период
- 9. Радиочувствительность клеток на разных стадиях клеточного цикла – эксперименты с синхронизированными клетками (остановка в одной из
- 10. Радиационные эффекты, регистрируемые на уровне клетки
- 11. Радиационная задержка клеточного деления впервые количественно исследована в 50-е годы 20-го века 60-е гг. - опыты
- 12. Длительность задержки клеточного цикла Длительность задержки деления зависит от того, в какой фазе цикла произошло облучение
- 13. Судьба потомков облученной клетки Клетка облучена в S-фазе. 1 – погибшие клетки, 2 – гигантские клетки
- 14. Образование гигантских клеток Размеры могут в сотни раз превосходить нормальные (min в 15 раз по площади)
- 15. Утеря клоногенного потенциала (Clonogenic survival assay) основной способ оценки выживаемости клеток Синонимы: клоногенная активность, жизнеспособность, выживаемость
- 16. Влияние 241Am на жизнеспособность клеток E.coli работа выполнена в ИХБиФМ СО РАН Показано значительное ингибирующее действие
- 17. Токсическое влияние 241Am, на стационарную культуру клеток S. typhimurium ТА102. Показано повышение гибели бактериальных клеток в
- 18. Утеря клоногенного потенциала II. Оценка выживаемости клеток In vivo - Модификации метода для стволовых клеток (1961
- 19. Длительное сохранение повышенной гибели потомков (снижение клоногенной способности) сохранение гибели части потомков облученной клетки в течение
- 20. SOS-хромотест - метод анализа активности генотоксинов основан на индукции SOS ответа в клетках E. сoli (мутантах)
- 21. SOS-хромотест - метод анализа активности генотоксинов Анализ состоит в инкубации индикаторного штамма с увеличивающимися концентрациями тестируемых
- 22. Оценка генотоксичности 241Am в SOS-хромотесте Зарегистрирована достоверное увеличение активности β-галактозидазы (фактор индукции), которая коррелирует с индукцией
- 23. Клеточные изменения при апоптозе и некрозе Стадии апоптоза: http://www.biochemistry.ru/apoptos/apoptosis.html: 2) На ранних стадиях апоптоза, в отличии
- 24. Радиационная гибель клеток Апоптоз – (энергозависимый механизм) программируемая гибель клеток (ПГК). Годом признания апоптоза как физиологического
- 25. Радиационная гибель клеток – Некроз – «клеточная катастрофа» или вид ПГК (роль АТФ)?? Некроз реализуется при
- 26. Причины радиационной гибели клеток В результате лучевого поражения клетки гибнут как по апоптотическому, так и по
- 27. Интерфазная и митотическая гибель клеток
- 28. Интерфазная гибель клеток = Гибель неделящихся (нервные, мышечные) или медленно делящихся (печень) клеток при облучении изменения,
- 29. Митотическая (репродуктивная) гибель клеток = в основном, быстроделящихся Гибель в митозе обусловлена хромосомными аберрациями: Поврежденные хромосомы
- 30. Последствия радиационных повреждений ДНК: Эффекты, наблюдаемые в клетках Схема строения хромосомы в поздней профазе — метафазе
- 31. Фрагментация хромосом - Образование микроядер фрагменты хромосом образуются в результате разрывов ДНК, В интерфазе фрагменты формируют
- 32. Хромосомные аберрации (Отражают число разрывов ДНК и дефекты их репарации) метод широко используется для оценки поглощенной
- 33. Аберрации хромосом Аберрации хромосом возникают, когда клетка облучена в предсинтетической стадии или в S-периоде, но до
- 34. Аберрации хроматид Аберрации хроматид возникают, когда клетка облучена после завершения репликации всей ДНК или того участка,
- 35. Нарушение геометрии хромосом метафазная пластинка облученных лимфоцитов в, г - метафазная пластинка, содержащая дицентрические хромосомы, кольцевые
- 36. Транслокации Некоторые заболевания – следствия транслокаций: Рак : несколько форм рака являются следствием транслокаций - описано
- 37. Изучение транслокаций: методы дифференцированной окраски участков хромосом, присоединение флюоресцентной метки к фрагментам ДНК, комплементарных для ДНК
- 38. Стабильность аберраций Нестабильные аберрации – ацентрические фрагменты и дицентрики - ведут к гибели клетки или ее
- 39. Аберрации коррелируют с поглощенной дозой и используются для целей дозиметрии (подробно в след. лекциях): Аберрации лимфоцитов
- 40. Радиационно-индуцированная нестабильность генома Часть клеток, выживших после облучения, может давать функционально измененное потомство, в котором с
- 41. Нестабильность генома: Сохранение НГ на протяжении десятков поколений клеток животных было впервые показано при облучении в
- 42. Механизм индукции и поддержания генетической нестабильности мало изучены: В развитии генетической нестабильности играет роль: увеличение поражения
- 43. Примеры: эксперименты по облучению цитоплазмы и ядер: облучение участков цитоплазмы ускоренными ядрами гелия: при прохождении 4
- 44. Результаты экспериментов и выводы Облучение цитоплазмы приводит к учащению точковых мутаций (характерных для спонтанного мутагенеза) Локальное
- 45. «Генетическая нестабильность обусловлена длительно сохраняющимся изменением функционирования клетки как целого, передаваемым потомству посредством эпигенетических механизмов» (подробности
- 47. Скачать презентацию

Стадии лучевого поражения клетки
Физическая (неспецифическая) (10-16-10-14 с) – поглощение, перераспределение и
Стадии лучевого поражения клетки
Физическая (неспецифическая) (10-16-10-14 с) – поглощение, перераспределение и

Стадии лучевого поражения клетки
Физико-химическая (10-13-10-10 с) – реакции заряженных и возбужденных
Стадии лучевого поражения клетки
Физико-химическая (10-13-10-10 с) – реакции заряженных и возбужденных

Стадии лучевого поражения клетки
Химическая (10-7-10-6 с) – к этому времени уже
Стадии лучевого поражения клетки
Химическая (10-7-10-6 с) – к этому времени уже
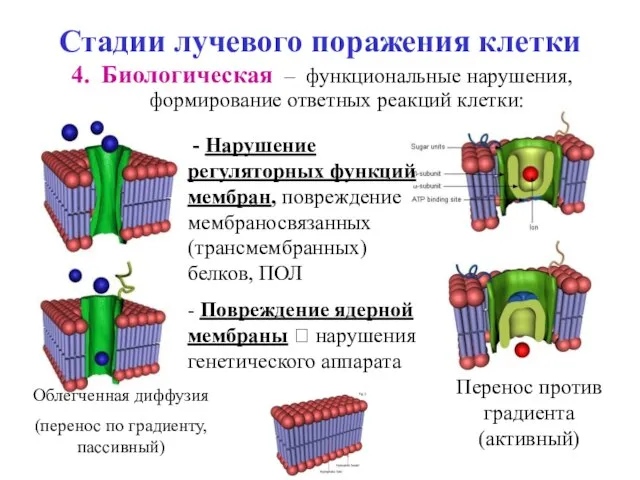
Стадии лучевого поражения клетки
Биологическая – функциональные нарушения, формирование ответных реакций клетки:
Перенос
Стадии лучевого поражения клетки
Биологическая – функциональные нарушения, формирование ответных реакций клетки:
Перенос

Биологическая стадия (продолжение)
Структурные повреждения нуклеиновых кислот приводят к нарушению процессов репликации,
Биологическая стадия (продолжение)
Структурные повреждения нуклеиновых кислот приводят к нарушению процессов репликации,
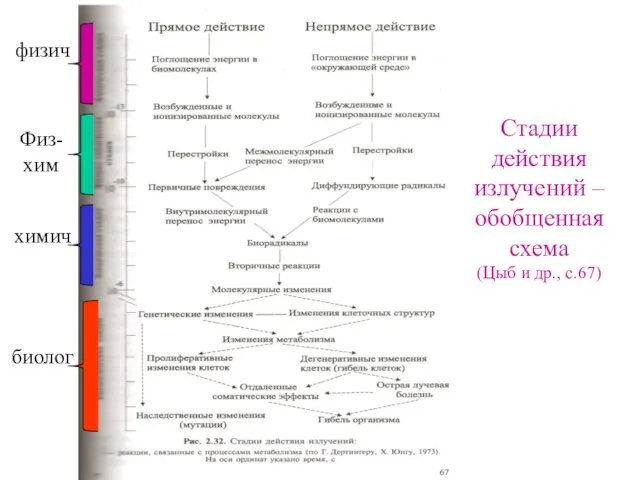
Стадии действия излучений – обобщенная схема
(Цыб и др., с.67)
физич
Физ-хим
химич
биолог
Стадии действия излучений – обобщенная схема
(Цыб и др., с.67)
физич
Физ-хим
химич
биолог
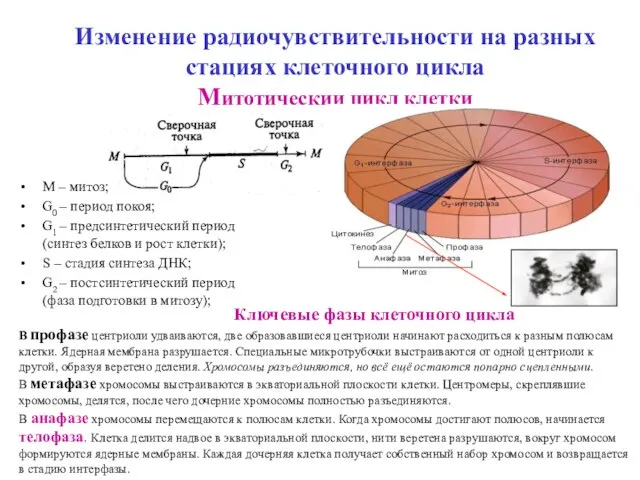
Изменение радиочувствительности на разных стациях клеточного цикла
Митотичеcкии цикл клетки
М
Изменение радиочувствительности на разных стациях клеточного цикла
Митотичеcкии цикл клетки
М
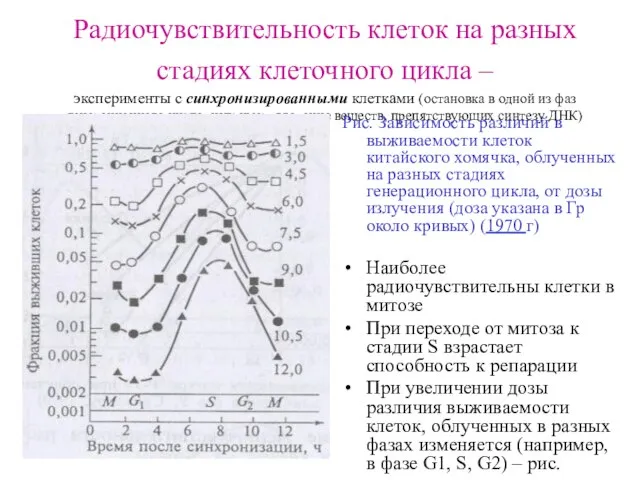
Радиочувствительность клеток на разных стадиях клеточного цикла –
эксперименты с синхронизированными
Радиочувствительность клеток на разных стадиях клеточного цикла – эксперименты с синхронизированными

Радиационные эффекты, регистрируемые на уровне клетки
Радиационные эффекты, регистрируемые на уровне клетки
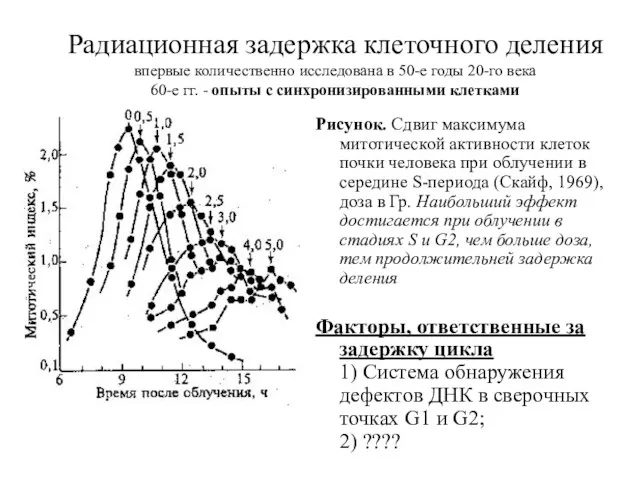
Радиационная задержка клеточного деления
впервые количественно исследована в 50-е годы 20-го века
60-е
Радиационная задержка клеточного деления впервые количественно исследована в 50-е годы 20-го века 60-е

Длительность задержки клеточного цикла
Длительность задержки деления зависит от того, в какой
Длительность задержки клеточного цикла
Длительность задержки деления зависит от того, в какой
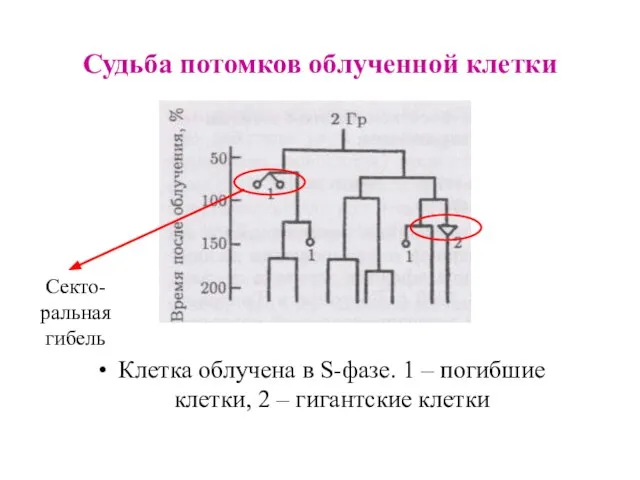
Судьба потомков облученной клетки
Клетка облучена в S-фазе. 1 – погибшие клетки,
Судьба потомков облученной клетки
Клетка облучена в S-фазе. 1 – погибшие клетки,

Образование гигантских клеток
Размеры могут в сотни раз превосходить нормальные (min в
Образование гигантских клеток
Размеры могут в сотни раз превосходить нормальные (min в

Утеря клоногенного потенциала
(Clonogenic survival assay)
основной способ оценки выживаемости клеток
Синонимы: клоногенная активность,
Утеря клоногенного потенциала (Clonogenic survival assay) основной способ оценки выживаемости клеток Синонимы: клоногенная активность,
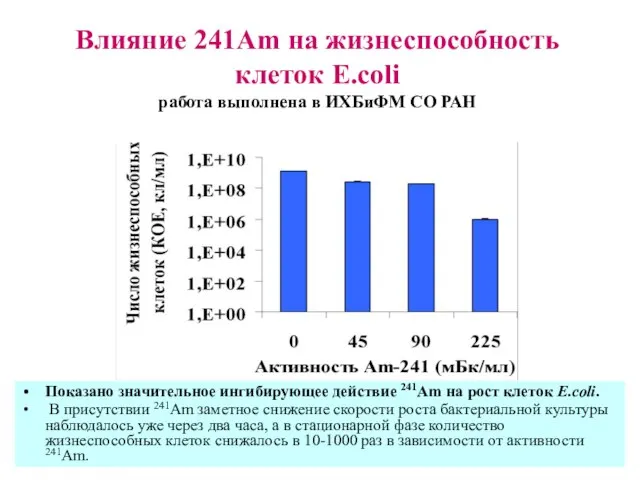
Влияние 241Am на жизнеспособность клеток E.coli
работа выполнена в ИХБиФМ СО РАН
Показано
Влияние 241Am на жизнеспособность клеток E.coli
работа выполнена в ИХБиФМ СО РАН
Показано

Токсическое влияние 241Am, на стационарную культуру клеток S. typhimurium ТА102.
Показано повышение
Токсическое влияние 241Am, на стационарную культуру клеток S. typhimurium ТА102.
Показано повышение

Утеря клоногенного потенциала
II. Оценка выживаемости клеток In vivo - Модификации
Утеря клоногенного потенциала
II. Оценка выживаемости клеток In vivo - Модификации
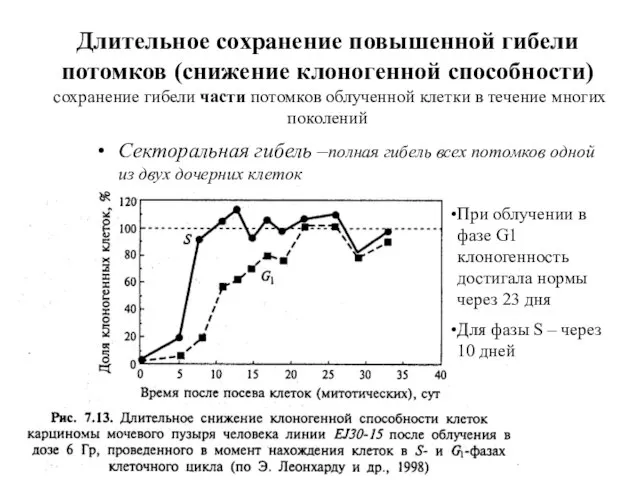
Длительное сохранение повышенной гибели потомков (снижение клоногенной способности)
сохранение гибели части
Длительное сохранение повышенной гибели потомков (снижение клоногенной способности) сохранение гибели части

SOS-хромотест - метод анализа активности генотоксинов
основан на индукции SOS ответа в
SOS-хромотест - метод анализа активности генотоксинов
основан на индукции SOS ответа в

SOS-хромотест - метод анализа активности генотоксинов
Анализ состоит в инкубации индикаторного штамма
SOS-хромотест - метод анализа активности генотоксинов
Анализ состоит в инкубации индикаторного штамма
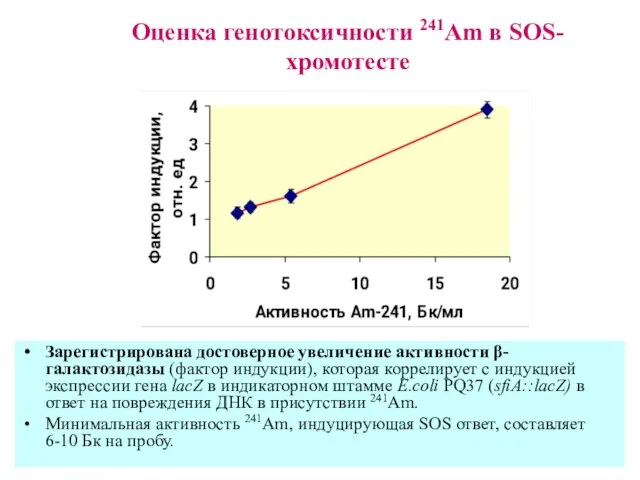
Оценка генотоксичности 241Am в SOS-хромотесте
Зарегистрирована достоверное увеличение активности β-галактозидазы (фактор индукции),
Оценка генотоксичности 241Am в SOS-хромотесте
Зарегистрирована достоверное увеличение активности β-галактозидазы (фактор индукции),

Клеточные изменения при апоптозе и некрозе
Стадии апоптоза:
http://www.biochemistry.ru/apoptos/apoptosis.html:
2) На ранних стадиях
Клеточные изменения при апоптозе и некрозе
Стадии апоптоза:
http://www.biochemistry.ru/apoptos/apoptosis.html:
2) На ранних стадиях

Радиационная гибель клеток
Апоптоз – (энергозависимый механизм) программируемая гибель клеток (ПГК). Годом
Радиационная гибель клеток
Апоптоз – (энергозависимый механизм) программируемая гибель клеток (ПГК). Годом

Радиационная гибель клеток –
Некроз – «клеточная катастрофа» или вид ПГК
Радиационная гибель клеток –
Некроз – «клеточная катастрофа» или вид ПГК

Причины радиационной гибели клеток
В результате лучевого поражения клетки гибнут как
Причины радиационной гибели клеток
В результате лучевого поражения клетки гибнут как

Интерфазная и митотическая гибель клеток
Интерфазная и митотическая гибель клеток

Интерфазная гибель клеток = Гибель неделящихся (нервные, мышечные) или медленно делящихся
Интерфазная гибель клеток = Гибель неделящихся (нервные, мышечные) или медленно делящихся

Митотическая (репродуктивная) гибель клеток
= в основном, быстроделящихся
Гибель в митозе
Митотическая (репродуктивная) гибель клеток
= в основном, быстроделящихся
Гибель в митозе
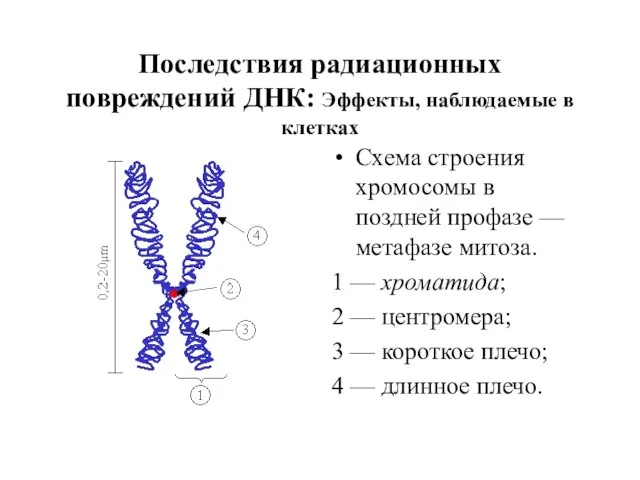
Последствия радиационных повреждений ДНК: Эффекты, наблюдаемые в клетках
Схема строения хромосомы в
Последствия радиационных повреждений ДНК: Эффекты, наблюдаемые в клетках
Схема строения хромосомы в

Фрагментация хромосом - Образование микроядер
фрагменты хромосом образуются в результате разрывов ДНК,
В
Фрагментация хромосом - Образование микроядер
фрагменты хромосом образуются в результате разрывов ДНК,
В
Хромосомные аберрации
(Отражают число разрывов ДНК и дефекты их репарации)
метод широко используется
Хромосомные аберрации (Отражают число разрывов ДНК и дефекты их репарации) метод широко используется

Аберрации хромосом
Аберрации хромосом возникают, когда клетка облучена в предсинтетической стадии или
Аберрации хромосом
Аберрации хромосом возникают, когда клетка облучена в предсинтетической стадии или

Аберрации хроматид
Аберрации хроматид возникают, когда клетка облучена после завершения репликации всей
Аберрации хроматид
Аберрации хроматид возникают, когда клетка облучена после завершения репликации всей
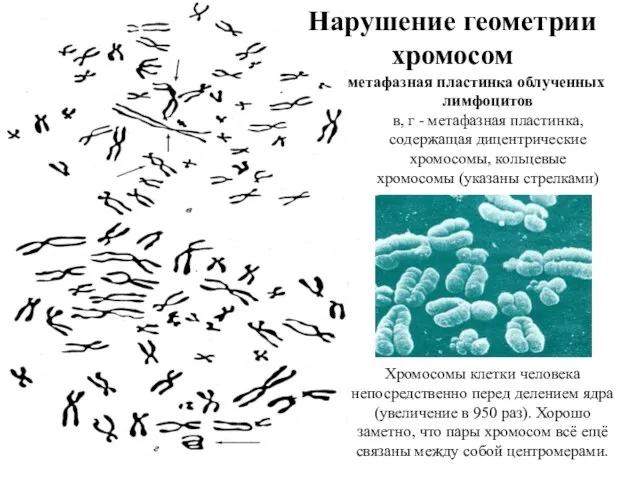
Нарушение геометрии хромосом
метафазная пластинка облученных лимфоцитов
в, г - метафазная пластинка, содержащая
Нарушение геометрии хромосом
метафазная пластинка облученных лимфоцитов в, г - метафазная пластинка, содержащая
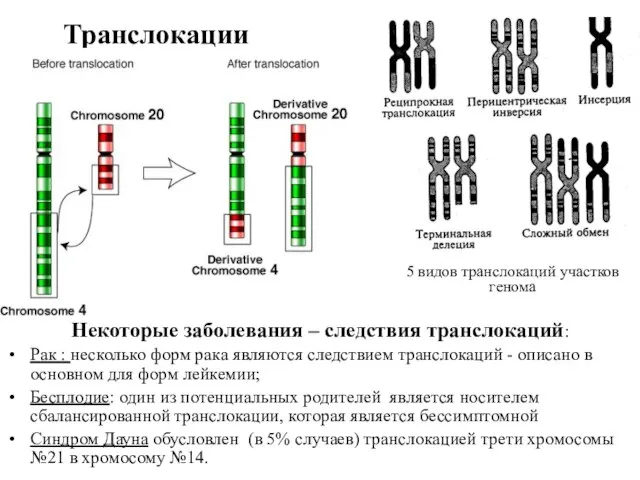
Транслокации
Некоторые заболевания – следствия транслокаций:
Рак : несколько форм рака являются следствием
Транслокации
Некоторые заболевания – следствия транслокаций:
Рак : несколько форм рака являются следствием
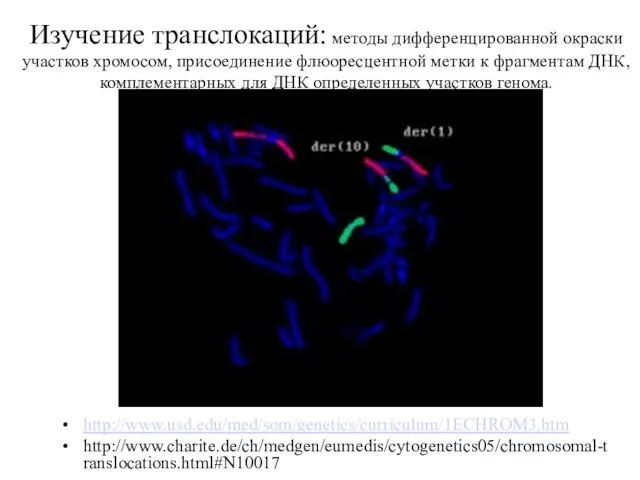
Изучение транслокаций: методы дифференцированной окраски участков хромосом, присоединение флюоресцентной метки к
Изучение транслокаций: методы дифференцированной окраски участков хромосом, присоединение флюоресцентной метки к

Стабильность аберраций
Нестабильные аберрации – ацентрические фрагменты и дицентрики - ведут к
Стабильность аберраций
Нестабильные аберрации – ацентрические фрагменты и дицентрики - ведут к

Аберрации коррелируют с поглощенной дозой и используются для целей дозиметрии (подробно
Аберрации коррелируют с поглощенной дозой и используются для целей дозиметрии (подробно

Радиационно-индуцированная нестабильность генома
Часть клеток, выживших после облучения, может давать функционально измененное
Радиационно-индуцированная нестабильность генома
Часть клеток, выживших после облучения, может давать функционально измененное

Нестабильность генома:
Сохранение НГ на протяжении десятков поколений клеток животных было впервые
Нестабильность генома:
Сохранение НГ на протяжении десятков поколений клеток животных было впервые

Механизм индукции и поддержания генетической нестабильности мало изучены:
В развитии генетической нестабильности
Механизм индукции и поддержания генетической нестабильности мало изучены:
В развитии генетической нестабильности

Примеры: эксперименты по облучению
цитоплазмы и ядер:
облучение участков цитоплазмы ускоренными ядрами
Примеры: эксперименты по облучению
цитоплазмы и ядер:
облучение участков цитоплазмы ускоренными ядрами

Результаты экспериментов и выводы
Облучение цитоплазмы приводит к учащению точковых мутаций (характерных
Результаты экспериментов и выводы
Облучение цитоплазмы приводит к учащению точковых мутаций (характерных

«Генетическая нестабильность обусловлена длительно сохраняющимся изменением функционирования клетки как целого, передаваемым
«Генетическая нестабильность обусловлена длительно сохраняющимся изменением функционирования клетки как целого, передаваемым
 Логопедический массаж
Логопедический массаж Кишечные инфекции. Сепсис
Кишечные инфекции. Сепсис Правильная осанка - залог здоровья
Правильная осанка - залог здоровья Этиология и патогенез болезней пародонта
Этиология и патогенез болезней пародонта Портальная гипертензия
Портальная гипертензия Применение микротоковой терапии в медицине и косметологии
Применение микротоковой терапии в медицине и косметологии Ветряная оспа, клиника, диагностика и лечение
Ветряная оспа, клиника, диагностика и лечение Рекреационная деятельность
Рекреационная деятельность Рахит. Основные причины дефицита фосфатов и солей кальция у детей раннего возраста
Рахит. Основные причины дефицита фосфатов и солей кальция у детей раннего возраста Синдром Лайелла
Синдром Лайелла Травматология және ортопедиядағы консервативты емдеу әдістері
Травматология және ортопедиядағы консервативты емдеу әдістері Инфекционный бронхит кур
Инфекционный бронхит кур СРС: Роль медико-генетической консультации в профилактике и ранней диагностике аномалий развития плода
СРС: Роль медико-генетической консультации в профилактике и ранней диагностике аномалий развития плода Устройства для ВК доступа. Новые методики в анестезиологии реаниматологии
Устройства для ВК доступа. Новые методики в анестезиологии реаниматологии Анестезиология и реаниматология
Анестезиология и реаниматология Оценка лабораторной диагностики сибирской язвы в объектах окружающей среды
Оценка лабораторной диагностики сибирской язвы в объектах окружающей среды Санаторий и здоровье
Санаторий и здоровье Қызметкерлердің қорғану құралдары. Жұқпалы аурулардың таралуын алдын алу (ВИЧ-жұқпасы), В-гепатиті және
Қызметкерлердің қорғану құралдары. Жұқпалы аурулардың таралуын алдын алу (ВИЧ-жұқпасы), В-гепатиті және Холера: этиология, эпидемиология, патогенез
Холера: этиология, эпидемиология, патогенез Тұқым қуалайтын белгілер мен аурулар
Тұқым қуалайтын белгілер мен аурулар Перенашивание беременности
Перенашивание беременности Понятие о психике и ее эволюция
Понятие о психике и ее эволюция Системная склеродермия
Системная склеродермия Синдром вегетативной дисфункции у детей и подростков
Синдром вегетативной дисфункции у детей и подростков Отравление рыбой
Отравление рыбой Изоантигенная несовместимость крови матери и плода
Изоантигенная несовместимость крови матери и плода Шкалы оценки тяжести травмы и тяжести состояния пострадавшего
Шкалы оценки тяжести травмы и тяжести состояния пострадавшего Медицинская информатика
Медицинская информатика