- Механизмы гормонального канцерогенеза

Содержание
- 2. Частота гормонозависимых форм рака у женщин в г. Новосибирске (на 100 000)
- 3. Факторы риска Поздняя менопауза Репродуктивные факторы Ожирение Бесплодие Увеличение уровня эстрогенов
- 4. Зависимость случаев гормоно-зависимых видов рака от возраста (n=3850)
- 5. Патогенез гормонального рака Карцинома Изменение внеклеточного матрикса Изменение клеточных сигналов Изменение рецепции эстрогенов Эстрогены, канцерогены Геномные
- 6. Концепция гормонального канцерогенеза Увеличение активности ароматазы Увеличение Уровня эстрогенов Снижение активности ферментов метаболизма Повышение активности сульфатазы
- 7. ПОДХОДЫ К ИЗУЧЕНИЮ ГОРМОНОЗАВИСИМЫХ ФОРМ РАКА Генетический анализ (полиморфизм генов) Молекулярная характеристика опухоли Технологии Omic (геномный,
- 8. Сигнальная трансдукция в гормональном канцерогенезе
- 9. Передача клеточного сигнала (Сигнальная трансдукция – СТ) дифференцировка пролиферация апоптоз межклеточные взаимодействия цитоскелет миграция энергетический метаболизм
- 10. Клетка постоянно получает сигналы из внеклеточного пространства, передает их и формирует адекватный ответ. СТ-механизм основан на
- 11. Характеристика RTKs У человека идентифицировано 58 RTKs, которые подразделяются на 20 подсемейств Все рецепторы имеют сходную
- 12. Семейство Epidermal growth factor receptor (EGFR) EGF был изолирован в1962 Stanley Cohen. Его взаимодействие с EGFR
- 13. Активируемые пути СТ
- 14. Нарушение метаболизма и рецепции эстрогенов
- 15. Нарушение метаболизма эстрогенов запускает гормональный канцерогенез, образование злокачественной опухоли в гормонозависимой ткани
- 16. Промоторный тип канцерогенеза Переэкспрессия ER-α в нормальной ткани увеличивает чувствительность к эстрогенам, повышает риск возникновения гормонозависимой
- 17. ER-опосредованные сигналы Доменная структура эстрогеновых рецепторов Эстрогены действуют через эстрогеновые рецепторы (ER, estrogen receptor). На настоящий
- 18. Структура эстрогенового рецептора
- 19. Стимуляция ER-сигнального пути эстрадиолом (Е2)
- 20. Модель действия эстрогенов ER NR-box NR-box Транскрипция генов Корегуляторы Факторы транскрипции Мишени в цитозоли MAPK PI3K/Akt
- 21. 2 ER ER ER ER P P TF P TF ER ER Протеин-киназный каскад eNOS P
- 22. Распределение эстрогеновых рецепторов в организме человека Центральная нервная система ERα, ERβ Печень ERα Кость ERα, ERβ
- 23. Строение ДНК-связывающего домена
- 24. Взаимодействие рецептора с ERE
- 25. Гены мишени эстрогеновых рецепторов Активация Супрессия Ген холин-ацетилтрансферазы Липопротеин липазный ген Ген фолликулостимулирующего гормона Ген хинолон
- 26. Схематическая иллюстрация регуляции транскрипции ERs в различных клетках
- 27. Научная проблема: Почему повышается количество ERальфа? Почему повышается его транскрипционная активность? Возможные причины: 1. Увеличение количества
- 28. ВЫКЛЮЧЕНИЕ ЯДЕРНЫХ РЕЦЕПТОРОВ
- 29. Загадки ГК: Сплайсированные варианты эстрогеновых рецепторов
- 30. Гипотетическая модель действия ERb на пути клеточной пролиферации
- 31. Схематическое изображение нарушения равновесия ERa и ERb при гормональном канцерогенезе
- 32. Аффинность лигандов к ERs (мкМ)
- 33. Генотоксический тип канцерогенеза Деградация эстрогенов в общих чертах представляет из себя деградацию ксенобиотиков – ключевую роль
- 34. Причины нарушения баланса Е2
- 35. Ксеноэстрогены и рецептор-опосредованный механизм
- 36. DDТ – потенциальный ксеноэстроген p,p’–DDT и о,p’-DDT in vitro связываются сAR. о,p’-DDТ in vitro связываются с
- 37. DDТ индуктор CYP2B в печени крыс
- 38. CYP1A1/2, CYP1B1 и CYP3A в метаболизме эстрогенов. Эффект ДДТ
- 39. Экспрессия CYP19 (Aromatase) A). Экспрессия CYP19 в матке и яичниках крыс, обработанных ДДТ B) Экспрессия CYP19
- 40. Экспрессия генов Erα и CyclinD1
- 41. Предполагаемый механизм действия DDT
- 42. МикроРНК в гормональном канцерогенезе МикроРНК- малые некодирующие РНК длиной 19-23 нуклеотида. Регулируют экспрессию более, чем 1/3
- 43. Эффекты DDT, BP, and MC на экспрессию miR-21/221/222/429 в печени(A) и яичников (B) самок крыс Вистар
- 44. Экспрессия miR в фиброаденоме МЖ и РМЖ
- 45. Профиль экспрессии МикроРНК в образцах РМЖ Без НАТ После НАТ
- 46. Таргетная гормональная терапия опухолей
- 47. Маркеры рака молочной железы
- 48. Маркеры рака эндометрия
- 49. Стратегия в лечении РМЖ
- 50. Гормональная терапия РМЖ Проблема: Поиск эффективных ингибиторов ERa (SERM)
- 52. Моделирование взаимодействия лигандов с ERs в экспериментах in vivo
- 53. Эксперименты in vivo с самками здоровых крыс Инъекции в течение 10 суток: 3 крысы – подсолнечное
- 54. Экспрессия генов cyclin D1 и ER-α в молочных железах крыс, обработанных эстрадиолом и тамоксифеном
- 55. Экспрессия генов cyclin D1 и ER-α в эндометрии крыс, обработанных эстрогеном и тамоксифеном
- 56. Экспрессия генов-«мишеней» в яичниках
- 57. Новые маркеры РМЖ для терапии
- 58. В опухолях молочной железы EGFR играет основную роль в усилении пролиферации и малигнизации Alvarez et al.
- 59. Резистентность к таргетам (Higgins and Baselga, 2011) Резистентность к анти- HER2 терапии может возникнуть, как результат
- 60. Роль BRCA1/2 в патогенезе РМЖ и рака яичников Наследственные формы рака молочной железы (около 5%) и
- 61. Онкосупрессор phosphatase and tensin homolog (PTEN)- маркер РЭ (Zhang and Yu, 2010) Потеря функции в 50%
- 62. Лечение РЭ ингибиторами mTOR
- 63. Лечение РЭ ингибиторами PI3K
- 65. Скачать презентацию
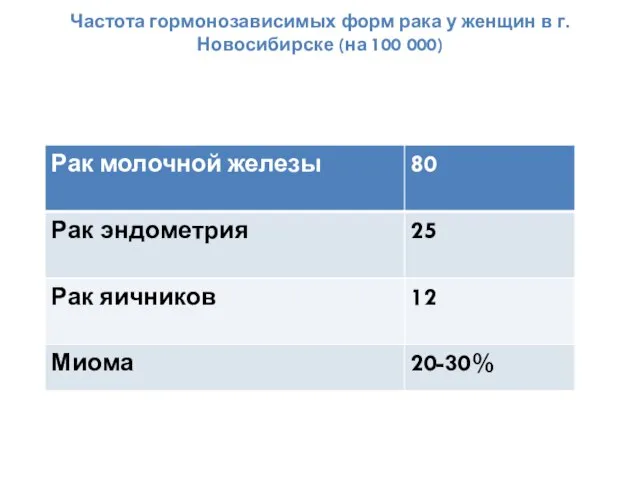
Частота гормонозависимых форм рака у женщин в г. Новосибирске (на 100
Частота гормонозависимых форм рака у женщин в г. Новосибирске (на 100
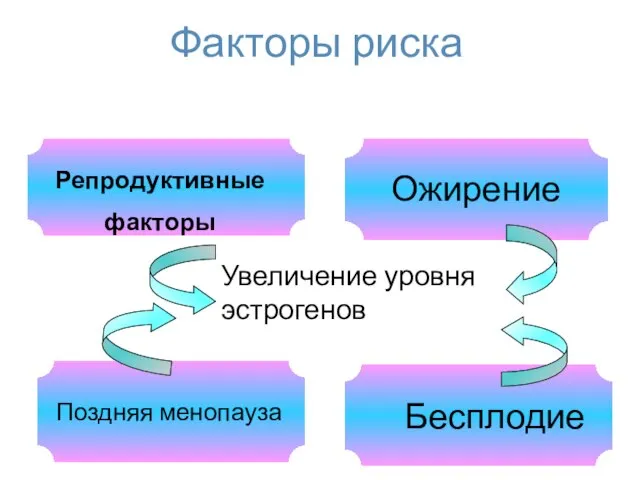
Факторы риска
Поздняя менопауза
Репродуктивные
факторы
Ожирение
Бесплодие
Увеличение уровня
эстрогенов
Факторы риска
Поздняя менопауза
Репродуктивные
факторы
Ожирение
Бесплодие
Увеличение уровня
эстрогенов
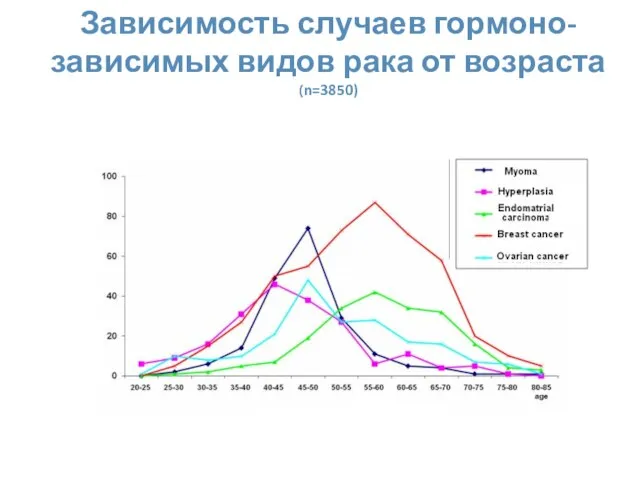
Зависимость случаев гормоно-зависимых видов рака от возраста (n=3850)
Зависимость случаев гормоно-зависимых видов рака от возраста (n=3850)
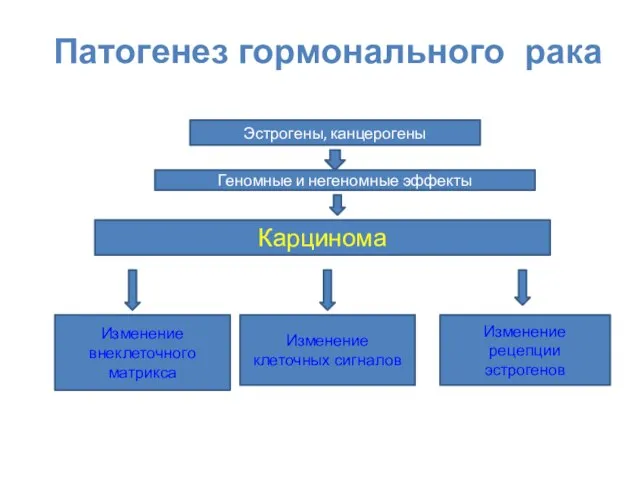
Патогенез гормонального рака
Карцинома
Изменение внеклеточного матрикса
Изменение клеточных сигналов
Изменение рецепции эстрогенов
Эстрогены, канцерогены
Геномные и
Патогенез гормонального рака
Карцинома
Изменение внеклеточного матрикса
Изменение клеточных сигналов
Изменение рецепции эстрогенов
Эстрогены, канцерогены
Геномные и
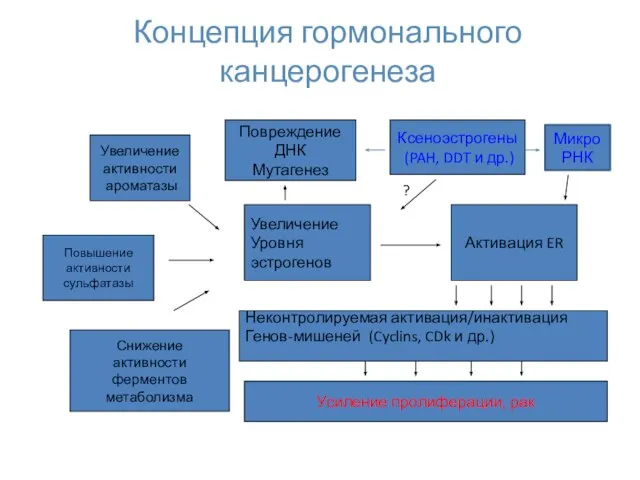
Концепция гормонального канцерогенеза
Увеличение
активности
ароматазы
Увеличение
Уровня
эстрогенов
Снижение
активности
ферментов
метаболизма
Повышение
активности
сульфатазы
Активация
Концепция гормонального канцерогенеза
Увеличение
активности
ароматазы
Увеличение
Уровня
эстрогенов
Снижение
активности
ферментов
метаболизма
Повышение
активности
сульфатазы
Активация
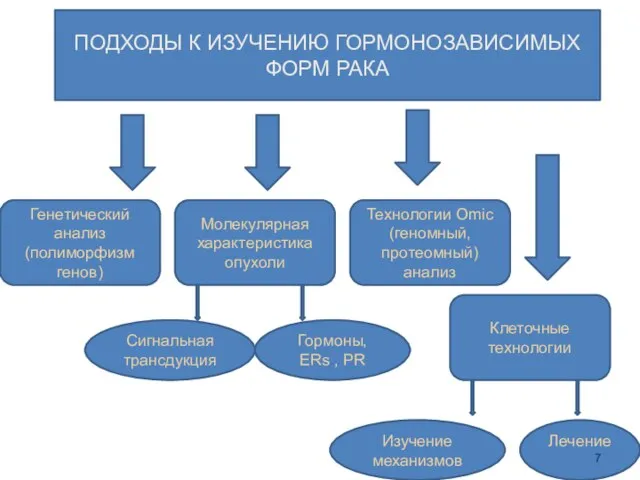
ПОДХОДЫ К ИЗУЧЕНИЮ ГОРМОНОЗАВИСИМЫХ ФОРМ РАКА
Генетический анализ (полиморфизм генов)
Молекулярная характеристика опухоли
Технологии
ПОДХОДЫ К ИЗУЧЕНИЮ ГОРМОНОЗАВИСИМЫХ ФОРМ РАКА
Генетический анализ (полиморфизм генов)
Молекулярная характеристика опухоли
Технологии

Сигнальная трансдукция в гормональном канцерогенезе
Сигнальная трансдукция в гормональном канцерогенезе
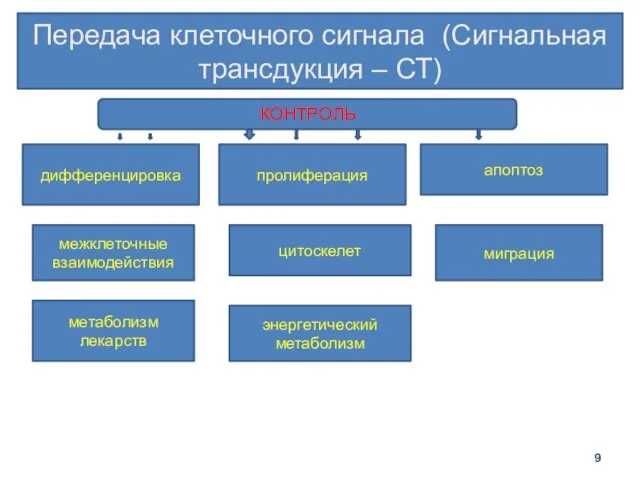
Передача клеточного сигнала (Сигнальная трансдукция – СТ)
дифференцировка
пролиферация
апоптоз
межклеточные взаимодействия
цитоскелет
миграция
энергетический
Передача клеточного сигнала (Сигнальная трансдукция – СТ)
дифференцировка
пролиферация
апоптоз
межклеточные взаимодействия
цитоскелет
миграция
энергетический
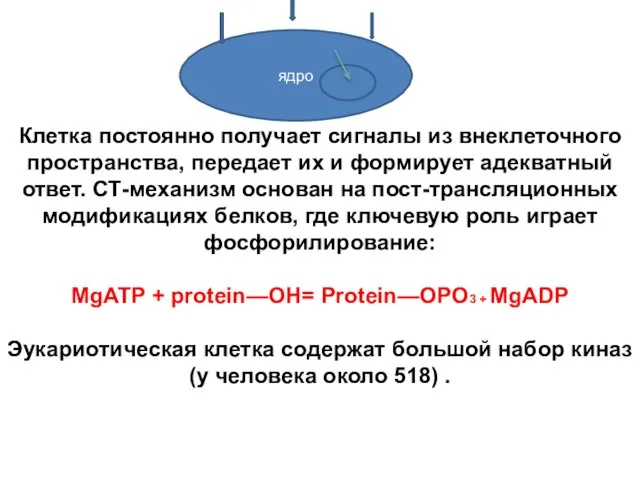
Клетка постоянно получает сигналы из внеклеточного пространства, передает их и формирует
Клетка постоянно получает сигналы из внеклеточного пространства, передает их и формирует

Характеристика RTKs
У человека идентифицировано 58 RTKs, которые подразделяются на 20 подсемейств
Все
Характеристика RTKs
У человека идентифицировано 58 RTKs, которые подразделяются на 20 подсемейств
Все

Семейство Epidermal growth factor receptor (EGFR)
EGF был изолирован в1962 Stanley Cohen.
Его
Семейство Epidermal growth factor receptor (EGFR)
EGF был изолирован в1962 Stanley Cohen.
Его
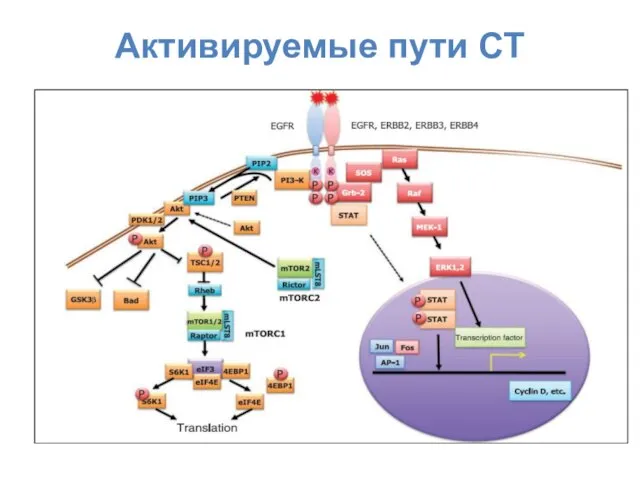
Активируемые пути СТ
Активируемые пути СТ

Нарушение метаболизма и рецепции эстрогенов
Нарушение метаболизма и рецепции эстрогенов
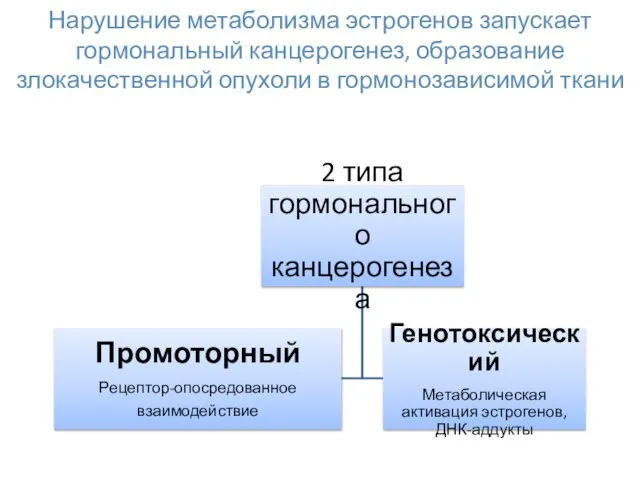
Нарушение метаболизма эстрогенов запускает гормональный канцерогенез, образование злокачественной опухоли в гормонозависимой
Нарушение метаболизма эстрогенов запускает гормональный канцерогенез, образование злокачественной опухоли в гормонозависимой
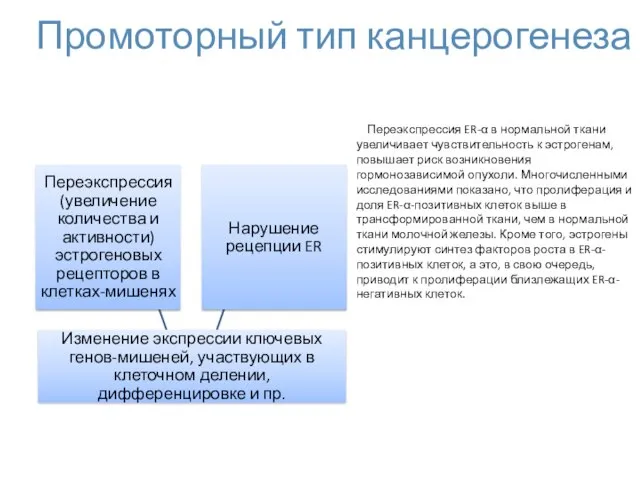
Промоторный тип канцерогенеза
Переэкспрессия ER-α в нормальной ткани увеличивает чувствительность к
Промоторный тип канцерогенеза
Переэкспрессия ER-α в нормальной ткани увеличивает чувствительность к
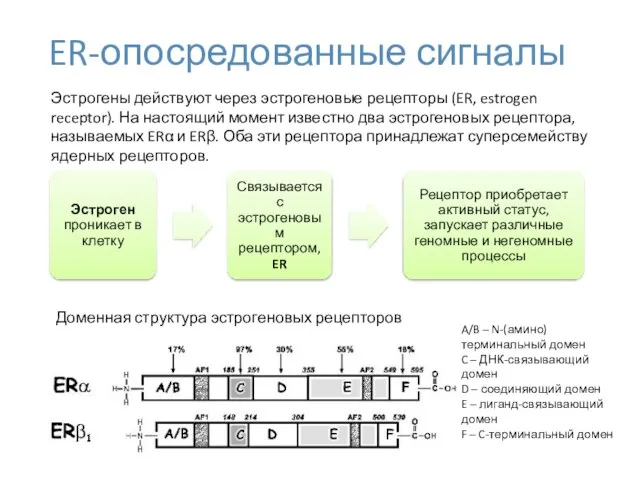
ER-опосредованные сигналы
Доменная структура эстрогеновых рецепторов
Эстрогены действуют через эстрогеновые рецепторы (ER, estrogen
ER-опосредованные сигналы
Доменная структура эстрогеновых рецепторов
Эстрогены действуют через эстрогеновые рецепторы (ER, estrogen
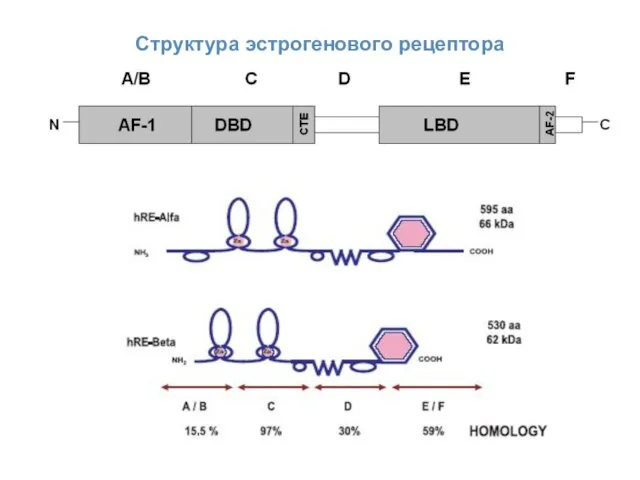
Структура эстрогенового рецептора
Структура эстрогенового рецептора
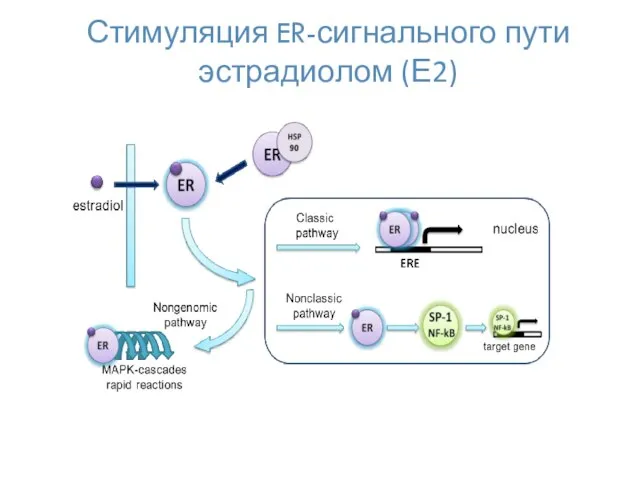
Стимуляция ER-сигнального пути эстрадиолом (Е2)
Стимуляция ER-сигнального пути эстрадиолом (Е2)
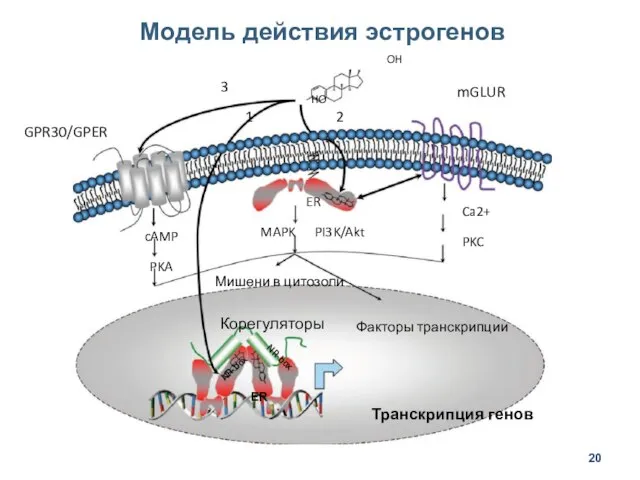
Модель действия эстрогенов
ER
NR-box
NR-box
Транскрипция генов
Корегуляторы
Факторы транскрипции
Мишени в цитозоли
MAPK PI3K/Akt
ER
Ca2+
PKC
cAMP
PKA
GPR30/GPER
mGLUR
OH
HO
1
2
3
Модель действия эстрогенов
ER
NR-box
NR-box
Транскрипция генов
Корегуляторы
Факторы транскрипции
Мишени в цитозоли
MAPK PI3K/Akt
ER
Ca2+
PKC
cAMP
PKA
GPR30/GPER
mGLUR
OH
HO
1
2
3
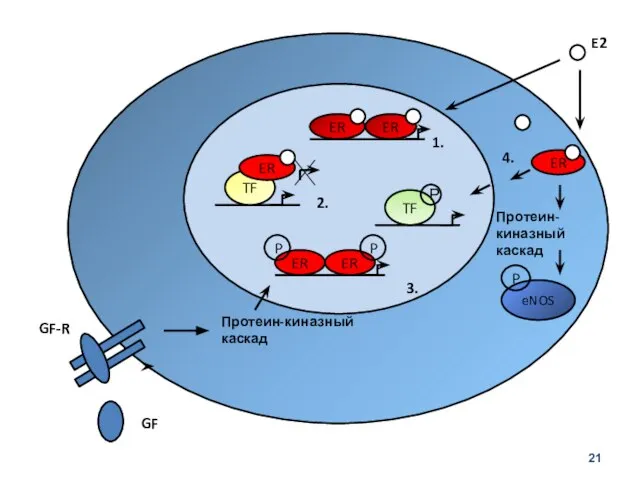
2
ER
ER
ER
ER
P
P
TF
P
TF
ER
ER
Протеин-киназный каскад
eNOS
P
E2
Протеин-киназный каскад
GF
GF-R
1.
2.
3.
4.
2
ER
ER
ER
ER
P
P
TF
P
TF
ER
ER
Протеин-киназный каскад
eNOS
P
E2
Протеин-киназный каскад
GF
GF-R
1.
2.
3.
4.

Распределение эстрогеновых рецепторов в организме человека
Центральная нервная система ERα, ERβ
Печень ERα
Кость
Распределение эстрогеновых рецепторов в организме человека
Центральная нервная система ERα, ERβ
Печень ERα
Кость
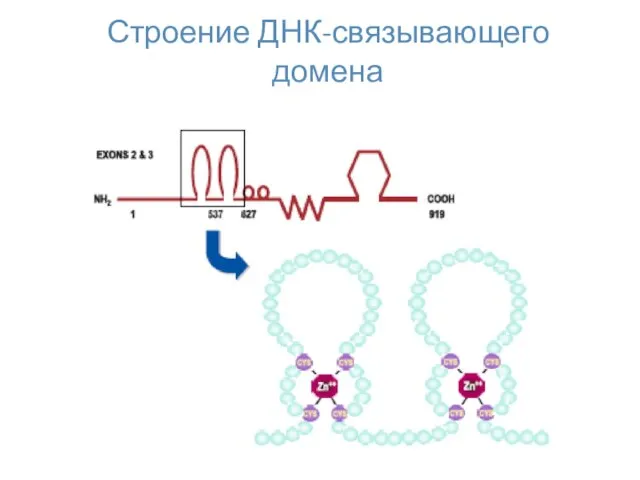
Строение ДНК-связывающего домена
Строение ДНК-связывающего домена
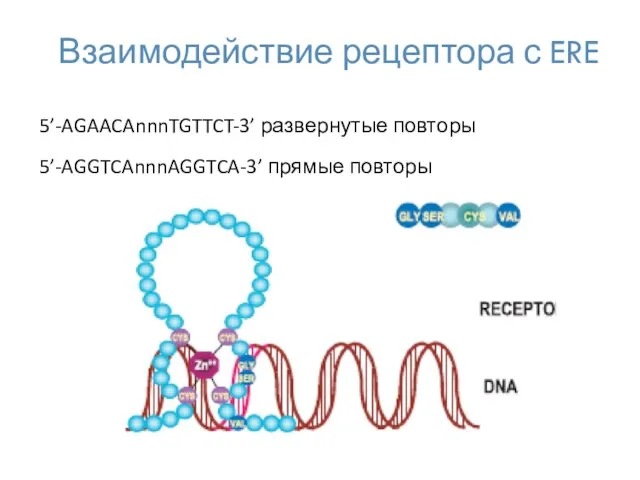
Взаимодействие рецептора с ERE
Взаимодействие рецептора с ERE

Гены мишени эстрогеновых рецепторов
Активация
Супрессия
Ген холин-ацетилтрансферазы
Липопротеин липазный ген
Ген
Гены мишени эстрогеновых рецепторов
Активация
Супрессия
Ген холин-ацетилтрансферазы
Липопротеин липазный ген
Ген

Схематическая иллюстрация регуляции транскрипции ERs в различных клетках
Схематическая иллюстрация регуляции транскрипции ERs в различных клетках

Научная проблема:
Почему повышается количество ERальфа?
Почему повышается его транскрипционная активность?
Возможные причины:
1.
Научная проблема:
Почему повышается количество ERальфа?
Почему повышается его транскрипционная активность?
Возможные причины:
1.

ВЫКЛЮЧЕНИЕ ЯДЕРНЫХ РЕЦЕПТОРОВ
ВЫКЛЮЧЕНИЕ ЯДЕРНЫХ РЕЦЕПТОРОВ
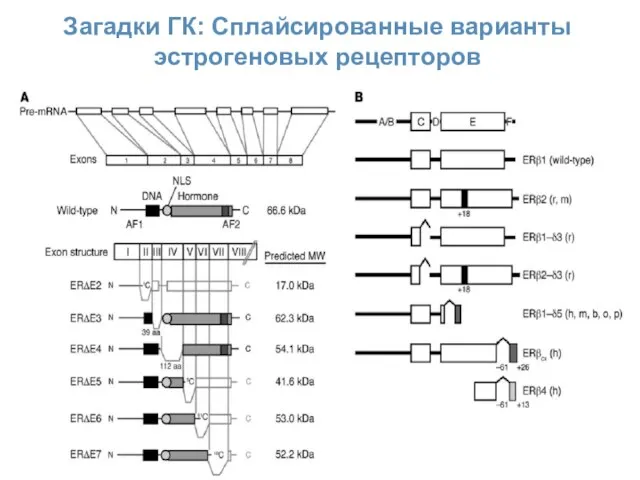
Загадки ГК: Сплайсированные варианты эстрогеновых рецепторов
Загадки ГК: Сплайсированные варианты эстрогеновых рецепторов
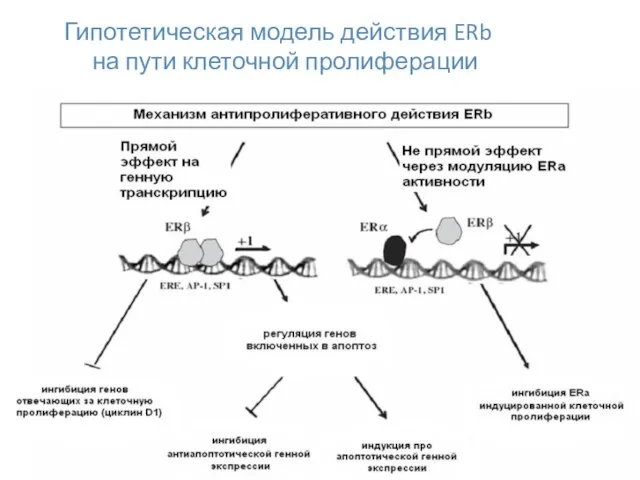
Гипотетическая модель действия ERb
на пути клеточной пролиферации
Гипотетическая модель действия ERb
на пути клеточной пролиферации
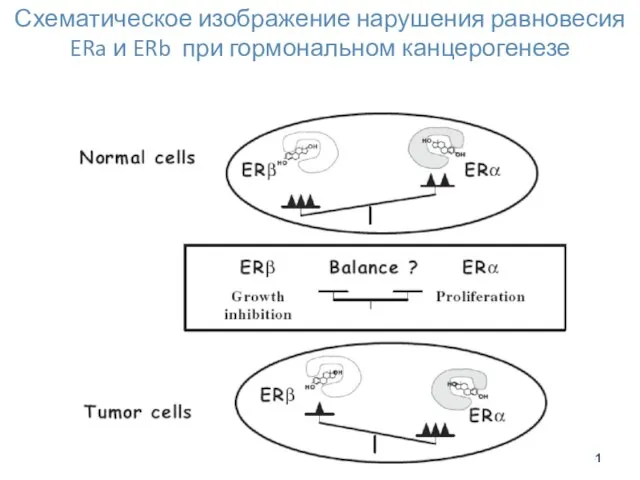
Схематическое изображение нарушения равновесия ERa и ERb при гормональном канцерогенезе
Схематическое изображение нарушения равновесия ERa и ERb при гормональном канцерогенезе
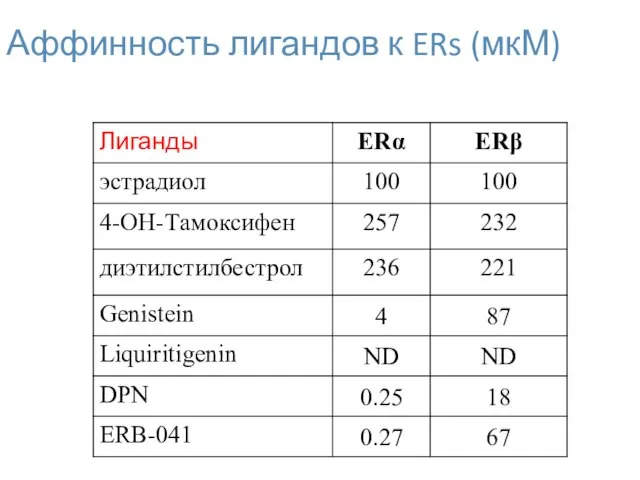
Аффинность лигандов к ERs (мкМ)
Аффинность лигандов к ERs (мкМ)
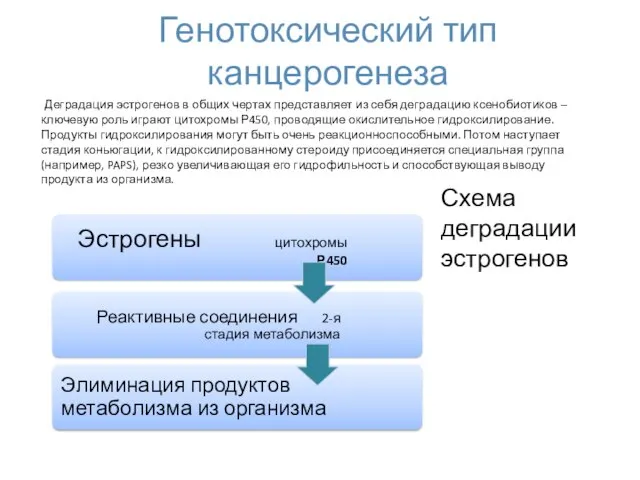
Генотоксический тип канцерогенеза
Деградация эстрогенов в общих чертах представляет из себя
Генотоксический тип канцерогенеза
Деградация эстрогенов в общих чертах представляет из себя

Причины нарушения баланса Е2
Причины нарушения баланса Е2

Ксеноэстрогены и рецептор-опосредованный механизм
Ксеноэстрогены и рецептор-опосредованный механизм
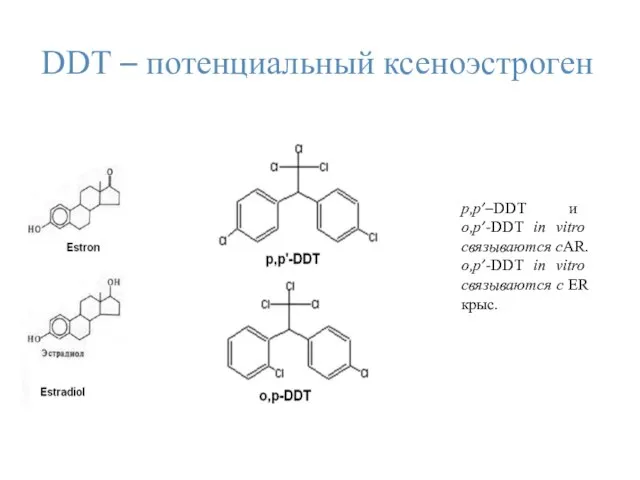
DDТ – потенциальный ксеноэстроген
p,p’–DDT и о,p’-DDT in vitro связываются сAR.
о,p’-DDТ in
DDТ – потенциальный ксеноэстроген
p,p’–DDT и о,p’-DDT in vitro связываются сAR.
о,p’-DDТ in
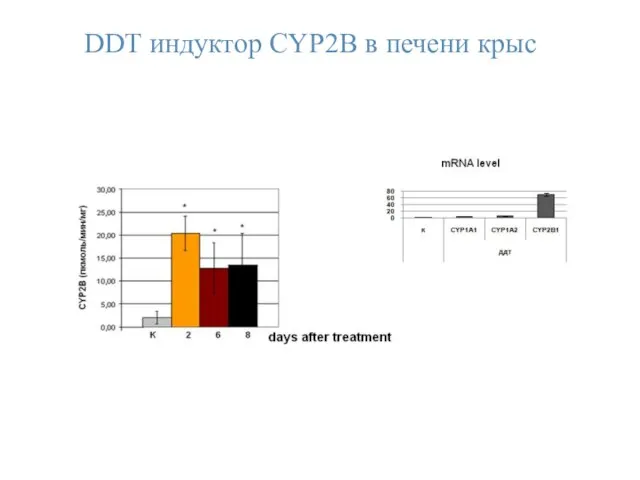
DDТ индуктор CYP2B в печени крыс
DDТ индуктор CYP2B в печени крыс
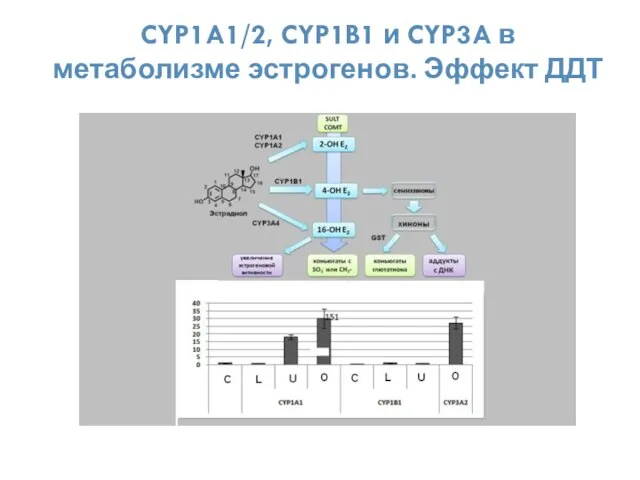
CYP1A1/2, CYP1B1 и CYP3A в метаболизме эстрогенов. Эффект ДДТ
CYP1A1/2, CYP1B1 и CYP3A в метаболизме эстрогенов. Эффект ДДТ
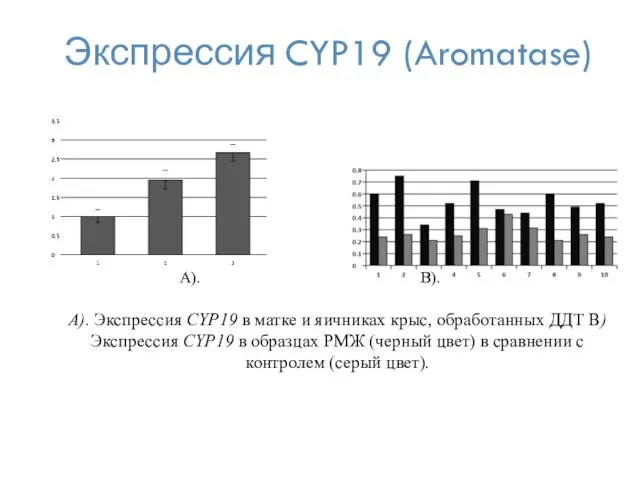
Экспрессия CYP19 (Aromatase)
A). Экспрессия CYP19 в матке и яичниках крыс, обработанных
Экспрессия CYP19 (Aromatase)
A). Экспрессия CYP19 в матке и яичниках крыс, обработанных
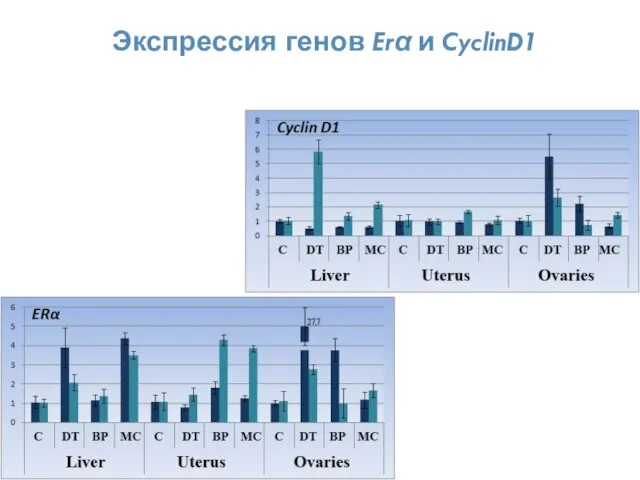
Экспрессия генов Erα и CyclinD1
Экспрессия генов Erα и CyclinD1
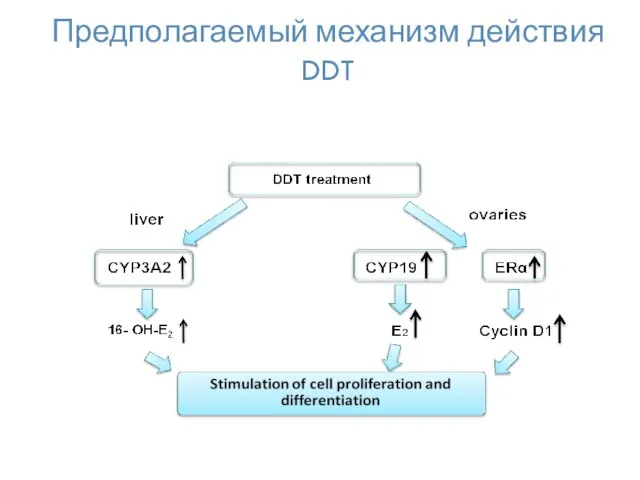
Предполагаемый механизм действия DDT
Предполагаемый механизм действия DDT
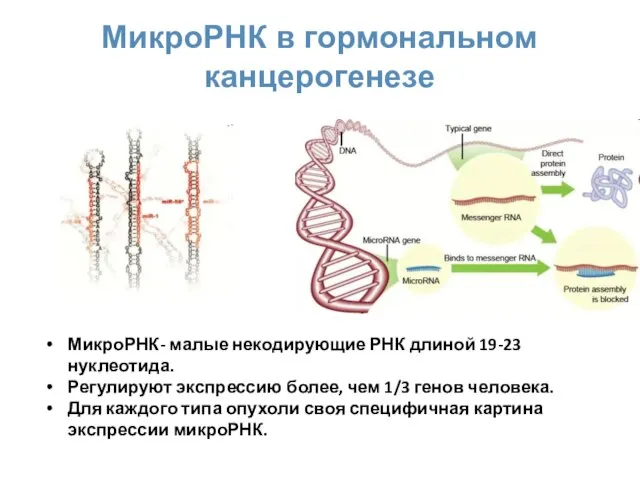
МикроРНК в гормональном канцерогенезе
МикроРНК- малые некодирующие РНК длиной 19-23 нуклеотида.
Регулируют экспрессию
МикроРНК в гормональном канцерогенезе
МикроРНК- малые некодирующие РНК длиной 19-23 нуклеотида.
Регулируют экспрессию

Эффекты DDT, BP, and MC на экспрессию miR-21/221/222/429 в печени(A) и
Эффекты DDT, BP, and MC на экспрессию miR-21/221/222/429 в печени(A) и

Экспрессия miR в фиброаденоме МЖ и РМЖ
Экспрессия miR в фиброаденоме МЖ и РМЖ
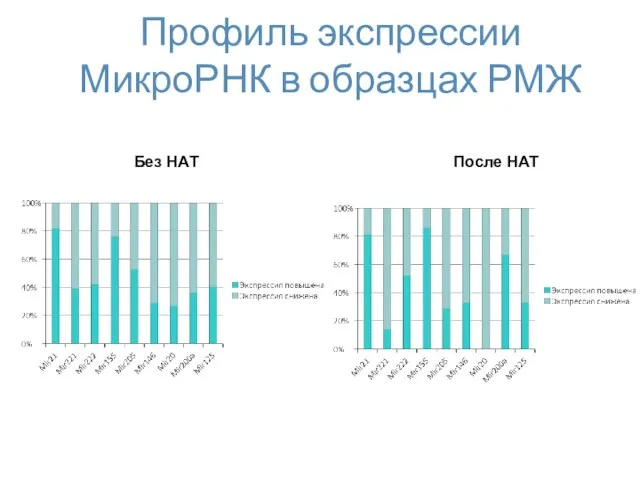
Профиль экспрессии МикроРНК в образцах РМЖ
Без НАТ
После НАТ
Профиль экспрессии МикроРНК в образцах РМЖ
Без НАТ
После НАТ

Таргетная гормональная терапия опухолей
Таргетная гормональная терапия опухолей

Маркеры рака молочной железы
Маркеры рака молочной железы
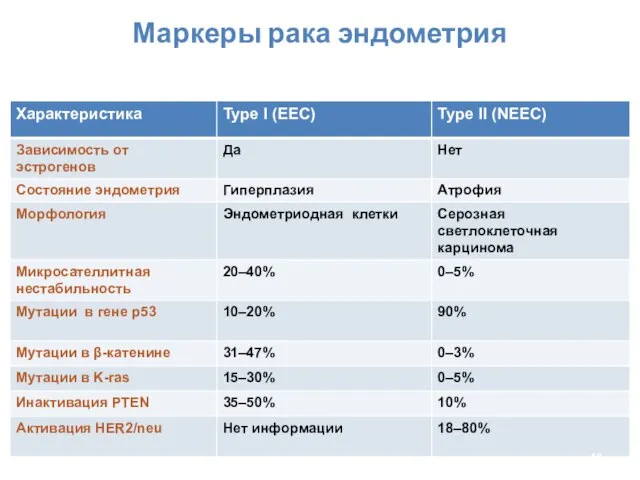
Маркеры рака эндометрия
Маркеры рака эндометрия

Стратегия в лечении РМЖ
Стратегия в лечении РМЖ

Гормональная терапия РМЖ
Проблема: Поиск эффективных ингибиторов ERa (SERM)
Гормональная терапия РМЖ
Проблема: Поиск эффективных ингибиторов ERa (SERM)


Моделирование взаимодействия лигандов с ERs в экспериментах in vivo
Моделирование взаимодействия лигандов с ERs в экспериментах in vivo
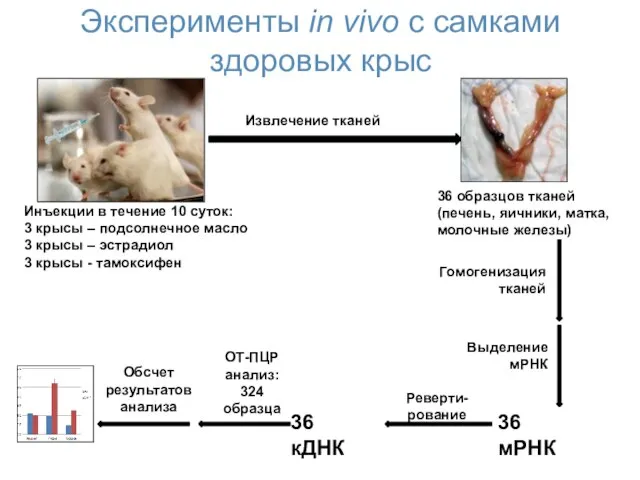
Эксперименты in vivo с самками здоровых крыс
Инъекции в течение 10
Эксперименты in vivo с самками здоровых крыс
Инъекции в течение 10
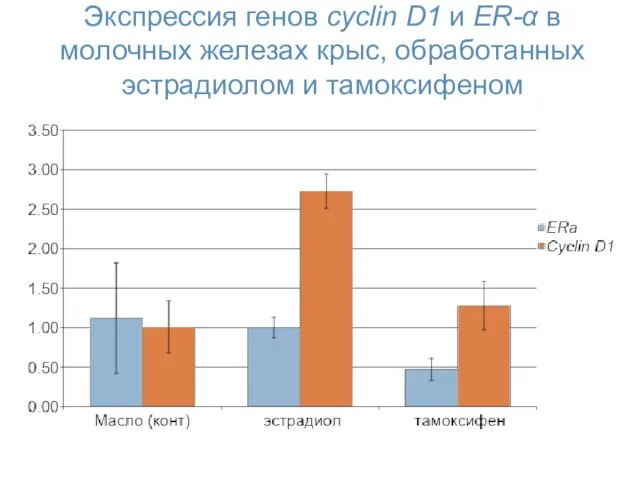
Экспрессия генов cyclin D1 и ER-α в молочных железах крыс, обработанных
Экспрессия генов cyclin D1 и ER-α в молочных железах крыс, обработанных
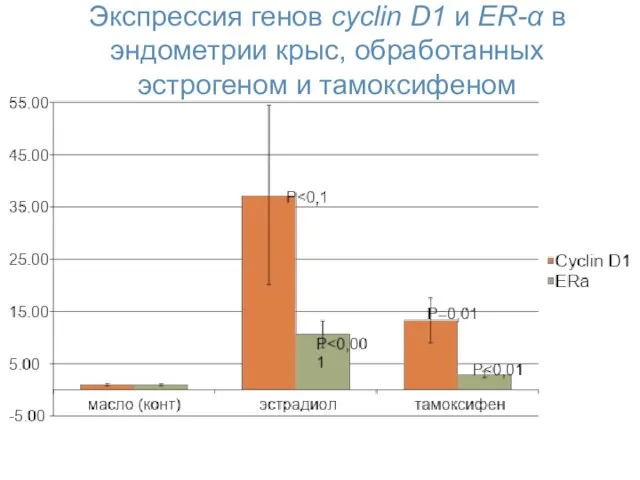
Экспрессия генов cyclin D1 и ER-α в эндометрии крыс, обработанных эстрогеном
Экспрессия генов cyclin D1 и ER-α в эндометрии крыс, обработанных эстрогеном
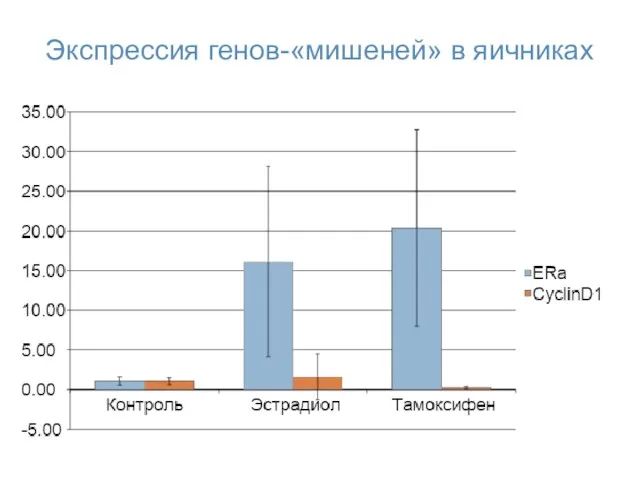
Экспрессия генов-«мишеней» в яичниках
Экспрессия генов-«мишеней» в яичниках
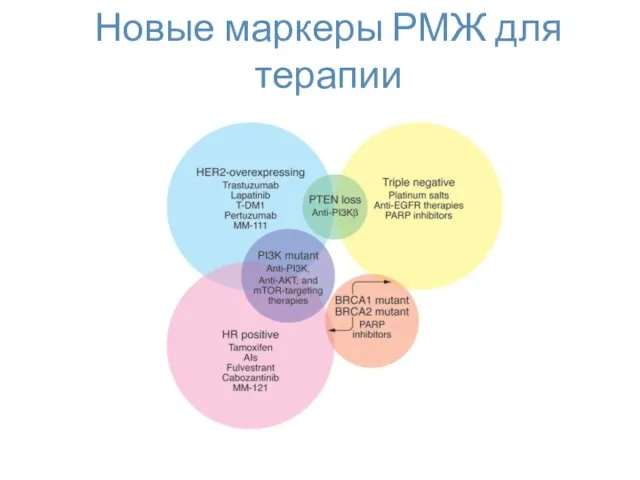
Новые маркеры РМЖ для терапии
Новые маркеры РМЖ для терапии

В опухолях молочной железы EGFR играет основную роль в усилении пролиферации
В опухолях молочной железы EGFR играет основную роль в усилении пролиферации
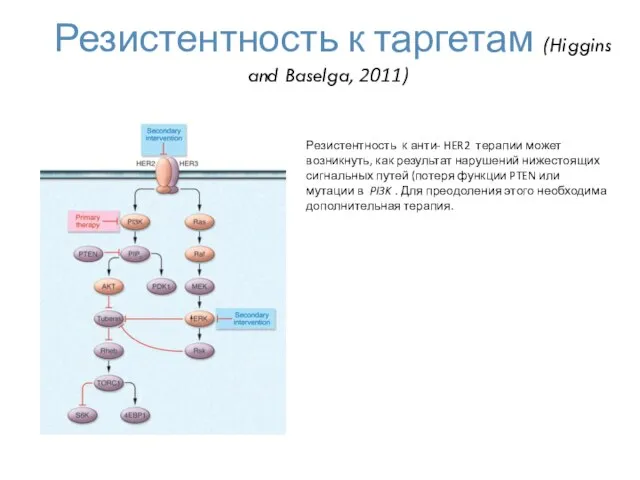
Резистентность к таргетам (Higgins and Baselga, 2011)
Резистентность к анти- HER2 терапии
Резистентность к таргетам (Higgins and Baselga, 2011)
Резистентность к анти- HER2 терапии

Роль BRCA1/2 в патогенезе РМЖ и рака яичников
Наследственные формы рака молочной
Роль BRCA1/2 в патогенезе РМЖ и рака яичников
Наследственные формы рака молочной
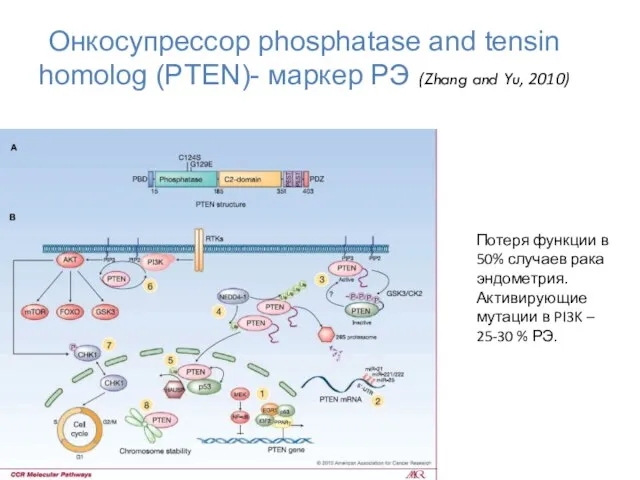
Онкосупрессор phosphatase and tensin homolog (PTEN)- маркер РЭ (Zhang and Yu,
Онкосупрессор phosphatase and tensin homolog (PTEN)- маркер РЭ (Zhang and Yu,
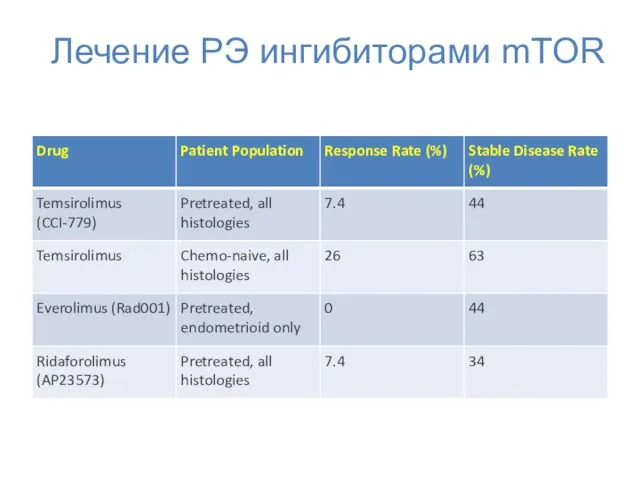
Лечение РЭ ингибиторами mTOR
Лечение РЭ ингибиторами mTOR
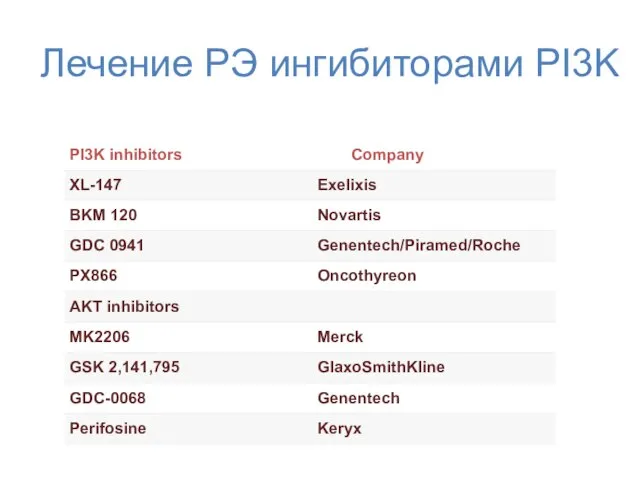
Лечение РЭ ингибиторами PI3K
Лечение РЭ ингибиторами PI3K
 Туберкулез. Терапия
Туберкулез. Терапия Теория развития интеллекта Жана Пиаже
Теория развития интеллекта Жана Пиаже Влияние метода обезболивания в родах на показатели КОС пуповинной крови новорожденных детей
Влияние метода обезболивания в родах на показатели КОС пуповинной крови новорожденных детей Младший школьный возраст от 6-7 до 10-11 лет
Младший школьный возраст от 6-7 до 10-11 лет Ромбоэмболия легочной артерии (ТЭЛА)
Ромбоэмболия легочной артерии (ТЭЛА) Хроническая венозная недостаточность и женщины
Хроническая венозная недостаточность и женщины Введение в дерматовенерологию. Значение дерматовенерологии для врачей гигиенистов. Основы диагностики заболеваний кожи
Введение в дерматовенерологию. Значение дерматовенерологии для врачей гигиенистов. Основы диагностики заболеваний кожи Туберкулезный спондилит
Туберкулезный спондилит Меланома. Причины возникновения меланомы
Меланома. Причины возникновения меланомы Психиатриядағы емдеу әдістері
Психиатриядағы емдеу әдістері Аускультация легких: побочные дыхательные шумы
Аускультация легких: побочные дыхательные шумы Эмпирические исследования познавательных процессов. ОПП- 2
Эмпирические исследования познавательных процессов. ОПП- 2 Жеке медициналық микробиологияның тапсырыс
Жеке медициналық микробиологияның тапсырыс Профілактика туберкульозу
Профілактика туберкульозу Қант диабетінің 1 типі кезінде ұзақ әсерлі инсулиннің әсерлігін анықтау
Қант диабетінің 1 типі кезінде ұзақ әсерлі инсулиннің әсерлігін анықтау Геморрагические диатезы
Геморрагические диатезы Системна склеродермія (системний склероз)
Системна склеродермія (системний склероз) Эндокринная система человека
Эндокринная система человека Паранеопластический артрит. Синдромная диагностика
Паранеопластический артрит. Синдромная диагностика Бауыр рагы
Бауыр рагы Митохондриальный геном и болезни человека
Митохондриальный геном и болезни человека Рак легень
Рак легень Анорексия-проблема нашего века
Анорексия-проблема нашего века Сердечно-легочная и церебральная реанимация догоспитальная и госпитальная (специализированная)
Сердечно-легочная и церебральная реанимация догоспитальная и госпитальная (специализированная) Физиотерапия в комплексном лечении болезней новорожденных
Физиотерапия в комплексном лечении болезней новорожденных Гемопоэз. Морфология клеточных элементов белой и красной крови. Приготовление мазка крови. Анемии. Занятия 3 и 4
Гемопоэз. Морфология клеточных элементов белой и красной крови. Приготовление мазка крови. Анемии. Занятия 3 и 4 Граднереллалар. Экологиясы. Адам үшін патогенділігі және науқас ағзасына орналасуы
Граднереллалар. Экологиясы. Адам үшін патогенділігі және науқас ағзасына орналасуы Современные исследованя органов зрения.Биомикроскопия. Эхоофтальмография .ФАГ. ОСТ. КТ. МРТ и их клинические особенности
Современные исследованя органов зрения.Биомикроскопия. Эхоофтальмография .ФАГ. ОСТ. КТ. МРТ и их клинические особенности