- Врожденный иммунитет. (Лекция 3)

Содержание
- 2. Врожденный иммунитет- наследственно закрепленная система защиты многоклеточных организмов от любых патогенных и непатогенных м/о, эндогенных продуктов
- 3. Врожденный иммунитет-первая линия защиты Миелоидные клетки Лимфоидные клетки Эпителиальные клетки и другие клетки НГ, МОН/МФ -фагоцитоз
- 4. Часть 1.Клеточные механизмы врожденного иммунитета
- 6. Эффекторные клетки врожденного иммунитета всегда готовы к осуществлению эффекторных функций (не требуется пролиферации, дифференцировки и межклеточных
- 7. ПАТТЕРН-РАСПОЗНАЮЩИЕ РЕЦЕПТОРЫ(PRR ) PAMP- патоген-ассоциированные молекулы –– консервативный рисунок аминокислотных последовательностей, характерный для большинства м/о и
- 8. Рецепторные молекулы миелоидных клеток Рецепторы обеспечивающие Распознавание -сигнальные (TLR1-11 и др) 3.Рецеторы опсонинов (Fc-рецепторы к Ig,
- 9. РЕГУЛЯЦИЯ ИММУНИТЕТА ЧЕРЕЗ СИГНАЛЬНЫЕ PRR ЭНДОЦИТОЗНЫЕ PRR (enPRR) РАМР enPRR Фагоцитоз и лизис Презентация антигена Адаптивный
- 10. Scavenger рецепторы Стрессорные молекулы – образы опасности DAMP (Danger-associatied molecular patterns) собственные молекулы организма, экспрессируемые на
- 11. главные клетки врожденного иммунитета- ФАГОЦИТЫ- микрофагоциты макрофагоциты НГ МОН/МФ Фагоцитоз- особый процесс поглощения клеткой ,крупных макромолекулярных
- 12. ФАГОЦИТОЗ –комплекс клеточных событий, в основе которых лежит распознавание, поглощение и элиминация из организма корпускулярных частиц,
- 14. Нейтрофильные гранулоциты Первыми (в течении 30 мин ) приходят в очаг воспаления, где проявляют разные виды
- 15. ПОВЕРХНОСТНЫЕ МЕМБРАННЫЕ РЕЦЕПТОРЫ НЕЙТРОФИЛЬНЫХ ГРАНУЛОЦИТОВ CR3(CD11b) CR1(CD35) C5aR LFA-1(CD11a) VLA-4(CD49d) FcγRIII (CD16) CD18 FcγRIIA(CD32) FcγRI(CD64)* CR4(CD11c)
- 16. (E.Kolaczkowska, P. Kubes Neutrophil recruitment and function in health and inflammation. Nature Reviews Immunology 13, 159-175
- 17. Распознавание чужеродных объектов НГ основано на рецепторном взаимодействии фагоцита и бактериальной клетки С этой целью используются
- 18. Поглощение и уничтожение микробов Взаимодействие НГ с опсонизируемой частицей инициирует процесс поглощения ее внутрь клетки Процесс
- 19. Специфические гранулы (вторичные) Лактоферин Лизоцим Цитохром b558 NADPH-оксидаза Коллагеназа Кателицидин Липокартин фосфолипаза CD11b/CD18 fMLP-R Азурофильные гранулы
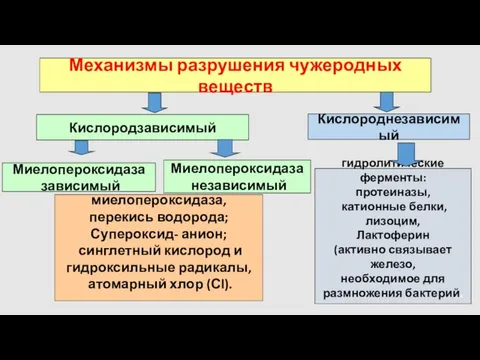
- 20. Механизмы разрушения чужеродных веществ Кислородзависимый Кислороднезависимый гидролитические ферменты: протеиназы, катионные белки, лизоцим, Лактоферин (активно связывает железо,
- 21. Кислород независимые факторы защиты Естественные полипептидные антибиотики- против Грамм(+), Грамм(-) бактерий, грибков и вирусов Основа литической
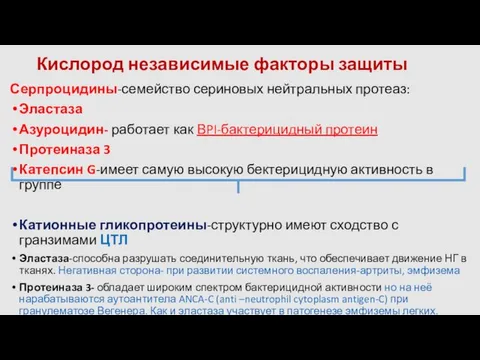
- 22. Кислород независимые факторы защиты Серпроцидины-семейство сериновых нейтральных протеаз: Эластаза Азуроцидин- работает как ВPI-бактерицидный протеин Протеиназа 3
- 23. Природные антибиотики-связующее звено между врожденным и адаптивным иммунитетом Дефензины НГ индуцируют миграцию МОН, ДК, наивных Тhо(CD4+CD45RA+)
- 24. Кислород независимые факторы защиты Лизоцим-(азурофильные и специфические гранулы НГ) расщепляет пиптидогликаны микробной клетки, хитиновый слой грибковых
- 25. Инициация фагоцитоза Запуск через Поступающий в клетку О2 не используется для энергии расходуется на образование бактерицидных
- 26. Респираторный взрыв Кислород- зависимая реакция-миелопероксидаза независимая
- 27. Респираторный взрыв Кислород-зависимая реакция с участием Миелопероксидазы (МП)-фермент азурофильных гранул, основная функция синтез высоко бактерицидных кислород
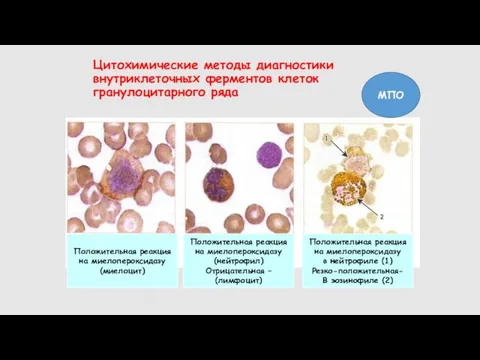
- 28. Цитохимические методы диагностики внутриклеточных ферментов клеток гранулоцитарного ряда Положительная реакция на миелопероксидазу (миелоцит) Положительная реакция на
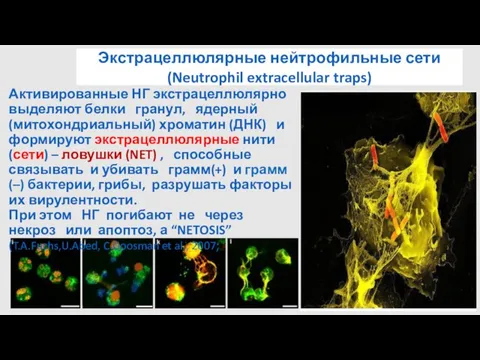
- 29. Экстрацеллюлярные нейтрофильные сети (Neutrophil extracellular traps) Активированные НГ экстрацеллюлярно выделяют белки гранул, ядерный (митохондриальный) хроматин (ДНК)
- 31. Дефекты в системе НГ Аллергические заболевания Персистирующая ВБИ респираторного тракта Аутоиммунные процессы Гнойно-септические заболевания Изолированные и
- 32. Макрофаги-ключевые клетки врожденного иммунитета Способны распознавать, поглощать и убивать инфицирующие агенты Очищать организм от собственных продуктов
- 34. Роль макрофага в иммунной системе:
- 35. Основная функция МФ-очищение организма от умирающих клеток-имеет выраженный противовоспалительный характер маркер CD14 scavenger рецепторы,Fc, манозо-связывающие рецепторы
- 36. Продукты секреции макрофагов
- 37. Различают МФ-резидентные и воспалительные Резидентные возникают при «плановой» миграции в ткани без воспаления Воспалительные-в процессе экстренной
- 38. Миграционная и хемотаксическая функция МФ такая же как у НГ хемоаттрактанты фрагменты системы комплемента, глобулины сыворотки
- 39. Способность одномоментно производить реактогенные метаболиты азота и кислорода существенно повышают бактерицидный потенциал МФ
- 40. NO зависимый киллинг Обладают микробицидной активностью: NO пероксинитрит(OONOI) (взаимодействие NO c супероксидным радикалом) NO2-,OH+
- 41. Дендритные клетки-профессиональные АПК Превосходят МФ по экспрессии HLA II класса Имеют костномозговое происхождение В периферической крови
- 43. ЭОЗИНОФИЛЫ, ТУЧНЫЕ КЛЕТКИ ,БАЗОФИЛЫ Основные эффекторы антипаразитарного иммунитета и аллергических реакций-открыл Пауль Эрлих Их объединяет: наличие
- 44. эозинофилы
- 45. имеют общий миелоидный предшественник локализуются: в подслизистом слое слизистых оболочек, соединительнотканном слое кожи, в серозных оболочках,
- 46. Естественные киллеры (NK-клетки, от nature killer) циркулирующие в крови CD56dim(мало)/CD16+ (Re для Fc-фрагмента IgG). популяция лимфоцитов,
- 47. NK-клетка Фенотип CD56 dimСD16++ Имеют Fc рецептор к IgG Распознает цели покрытые антителом Киллинг посредством антителозависимым
- 49. Активированные NK клетки цитокинами IL2 IFN IFN IL2 Главная физиологическая функция – киллерная в отношении активированных
- 50. Другие рецепторы естественных киллеров сложная система рецепторов распознающих -молекулы собственных клеток организма стресс-индуцированные клеточные лиганды, которые
- 51. Активирующий и ингибирующий сигналы NK. система регуляторных Re (inhibitory NK cell receptors). 2 больших семейства: KLRs
- 52. NKT - лимфоциты «Неклассическая» субпопуляция Т-лимфоцитов врожденного иммунитета сочетает признаки Т-л и NK, экспрессируют одновременно ТСR-СD3+
- 53. yδT – лимфоциты В ранний период после рождения, выполняют важные иммунорегуляторные функции при вирусных инфекциях. 2
- 54. Часть 2.ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА
- 55. СИСТЕМА КОМПЛЕМЕНТА (С)- -КОМПЛЕКС белков комплемента, их расщепленных фрагментов, рецепторов к компонентам комплемента, а также белков,
- 56. Белки системы комплемента представлены сывороточными белками (> 30), циркулирующими в кровеносном русле и присутствующими на мембранах
- 57. КОМПОНЕНТЫ КОМПЛЕМЕНТА СИНТЕЗИРУЮТСЯ: в печени- гепатоцитами (90%) макрофагами эпителием кишечника, почечных канальцев клетками эндотелия, кератинобластами, фибробластами
- 58. Пути активации комплемента Классический путь Альтернативный путь Лектиновый путь
- 59. КОМПОНЕНТЫ КЛАССИЧЕСКОГО ПУТИ C4 C2 C3 C1 комплекс
- 60. C4 КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ C3-КОНВЕРТАЗЫ Активация системы комплемента по классическому пути- индуцируется иммунными комплексами АГ-АТ
- 61. КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ C3-КОНВЕРТАЗЫ C4b Mg++ C4a C2 C4b2a - C3-конвертаза классического пути
- 62. КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ C5-КОНВЕРТАЗЫ C4b Mg++ C4a C2b C3 C4b2a3b -C5 конвертаза; запускает сборку МАК
- 63. Компоненты литического комплекса C6 C9 C5 C9 C9 C9
- 64. Активация C5 C5
- 65. Сборка литического комплекса МАК C6
- 66. Действие МАК на клетку мишень C 9 C 9 C 9 C 9 C 9 C
- 67. Лектиновый путь C4 C2 C4b2a - C3 конвертаза; активирует образование C5 -конвертазы MASP1 MASP2 Активация -индуцируется
- 68. Активация C5 C5
- 69. Сборка литического комплекса МАК C6
- 70. Действие МАК на клетку мишень C 9 C 9 C 9 C 9 C 9 C
- 71. Компоненты альтернативного пути C3 B D P Запуск начинается с С3 Содержание С3 составляет ½ концентрации
- 72. Спонтанная активация C3 C3 H2O B D Образование C3 конвертазы C3iBb комплекс короткоживущий-нестабильный C3
- 73. B D Спонтанное -образование C3b не разрушается C3-активация и амплификация C3 Комплекс C3bBb очень не стабилен
- 74. C3a B D C3 C3-активация и амплификация Этот комплекс эффективно стабилизируют полисахариды, гликолипиды, гликопротеины поверхности микроорганизмов.
- 75. C3a C3a C3 B D C3-активация и амплификация образовавшийся комплекс приобретает свойства С5-конвертазы и запускает формирование
- 76. Активация C5 C5
- 77. Сборка литического комплекса МАК C6
- 78. Действие МАК на клетку мишень C 9 C 9 C 9 C 9 C 9 C
- 79. C5-конвертазы классического, лектинового и альтернативного путей C3b Bb C3b C5-конвертаза альтернативного пути C5-конвертаза классического и лектинового
- 81. Центральный этап каскада комплемента Все три пути активации С приводят к образованию С3-конвертазы (классический путь, лектиновый
- 83. C3b Опсонин- способствуют фагоцитозу бактерий, на поверхности которых идет активация системы комплемента C5a Сильнейший хемоаттрактант для
- 84. Биологические эффекты C5a Активация нейтрофилов Адгезия нейтрофилов Миграция и хемотаксис нейтрофилов 4.Активация моноцитов 5.Дегрануляция тучных клеток
- 85. Обсонизация и фагоцитоз комплемент бактерия Фагоцитирующие клетки обсонизация распознавание фагоцитоз
- 86. РЕЦЕПТОРЫ К КОМПОНЕНТАМ КОМПЛЕМЕНТА Эр Т-лф В-лФ НГ,МФ,ЭОЗ ДК CR1(СD35) опсонизация В-лф Тимоциты ДК CR2(СD21) для
- 87. РЕГУЛЯЦИЯ АКТИВНОСТИ КОМПЛЕМЕНТА Регуляторы – белки, подавляющие отдельные стадии каскада активации С, ограничивают действие С на
- 88. РЕГУЛЯЦИЯ АКТИВНОСТИ КОМПЛЕМЕНТА Растворимые: Ингибитор С1 - эстеразы (C1-INH или С1-inh) блокирует протеолиз С2 и С4
- 90. Нарушения в системе комплемента ГЕНЕТИЧЕСКИ-ДЕТЕРМИНИРОВАННЫЕ дефекты компонентов C1 - C4 дефекты любых компонентов проявляются аутоиммунными заболеваниями
- 91. Дефект компонента C3 проявляется, рецидивирующими бактериальными пневмониями, менингитами, перитонитами Streptococcus pneumoniae, Staphylococcus aureus, Neisseria meningitidis Дефекты
- 92. Дефицит C1-ингибитора: ангионевротический отек
- 93. гипокомплементемия Сниженная продукция компонентов комплемента Усиленное расходование компонентов комплемента Формирование иммунных комплексов системная красная волчанка, постинфекционный
- 94. -группа протеинов синтезируемая гепатоцитами. В ответ на стрессорные факторы (воспаление, малигнизация, травма, обширное оперативное вмешательство, инфаркт
- 95. Гуморальные факторы:Белки острой фазы воспаления. Пентраксины С-реактивный белок Сывороточный амилоид Р Под действием Провоспалительных цитокинов Пентраксин
- 96. С- реактивный белок (СРБ) В норме (≈ 1 мкг/мл), при воспалительных процессах быстро нарастает в 20-25
- 97. Маннозосвязывающий лектин – МСЛ (MBL)- относится к семейству коллектинов - подобен компоненту комплемента C1q. Исходная концентрация
- 98. Под влиянием патогенов МФ,НГ ,ЭК активируются, секретируют цитокины и хемокины Лейкоциты мигрируют через сосудистую стенку в
- 99. Фагоциты. В начале данного этапа их эффективность невелика: число НГ и МФ в месте внедрения недостаточно
- 101. Скачать презентацию

Врожденный иммунитет-
наследственно закрепленная система защиты многоклеточных организмов от любых патогенных и
Врожденный иммунитет-
наследственно закрепленная система защиты многоклеточных организмов от любых патогенных и
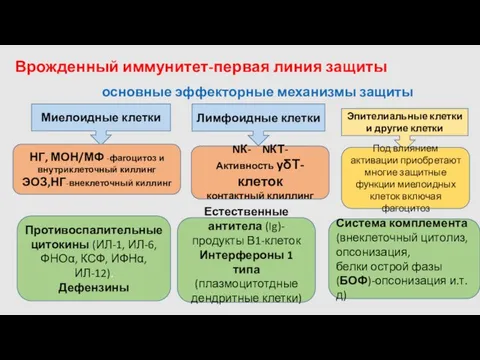
Врожденный иммунитет-первая линия защиты
Миелоидные клетки
Лимфоидные клетки
Эпителиальные клетки и другие клетки
НГ, МОН/МФ
Врожденный иммунитет-первая линия защиты
Миелоидные клетки
Лимфоидные клетки
Эпителиальные клетки и другие клетки
НГ, МОН/МФ
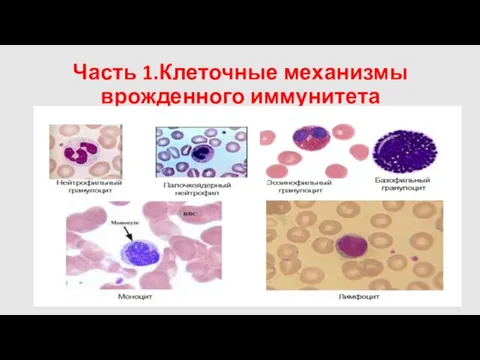
Часть 1.Клеточные механизмы врожденного иммунитета
Часть 1.Клеточные механизмы врожденного иммунитета
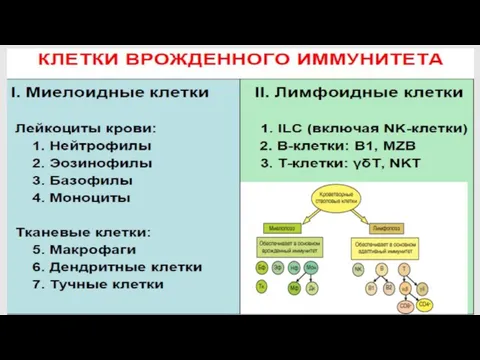

Эффекторные клетки врожденного иммунитета
всегда готовы к осуществлению эффекторных функций
(не требуется
Эффекторные клетки врожденного иммунитета
всегда готовы к осуществлению эффекторных функций
(не требуется

ПАТТЕРН-РАСПОЗНАЮЩИЕ РЕЦЕПТОРЫ(PRR )
PAMP- патоген-ассоциированные молекулы –– консервативный рисунок аминокислотных последовательностей, характерный
ПАТТЕРН-РАСПОЗНАЮЩИЕ РЕЦЕПТОРЫ(PRR )
PAMP- патоген-ассоциированные молекулы –– консервативный рисунок аминокислотных последовательностей, характерный

Рецепторные молекулы миелоидных клеток
Рецепторы обеспечивающие
Распознавание -сигнальные
(TLR1-11 и др)
3.Рецеторы опсонинов (Fc-рецепторы к
Рецепторные молекулы миелоидных клеток
Рецепторы обеспечивающие
Распознавание -сигнальные
(TLR1-11 и др)
3.Рецеторы опсонинов (Fc-рецепторы к
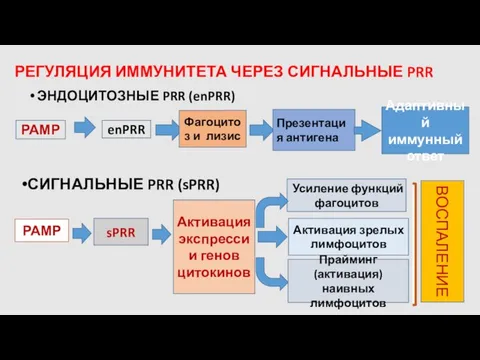
РЕГУЛЯЦИЯ ИММУНИТЕТА ЧЕРЕЗ СИГНАЛЬНЫЕ PRR
ЭНДОЦИТОЗНЫЕ PRR (enPRR)
РАМР
enPRR
Фагоцитоз и лизис
Презентация
РЕГУЛЯЦИЯ ИММУНИТЕТА ЧЕРЕЗ СИГНАЛЬНЫЕ PRR
ЭНДОЦИТОЗНЫЕ PRR (enPRR)
РАМР
enPRR
Фагоцитоз и лизис
Презентация

Scavenger рецепторы
Стрессорные молекулы – образы опасности DAMP (Danger-associatied molecular patterns)
собственные
Scavenger рецепторы
Стрессорные молекулы – образы опасности DAMP (Danger-associatied molecular patterns)
собственные
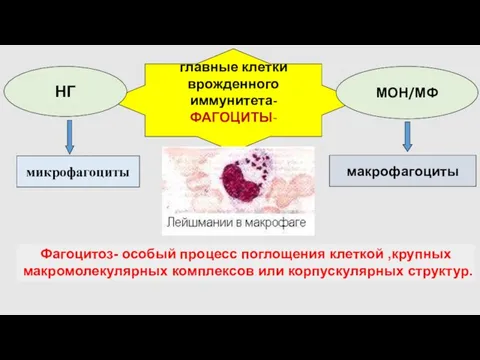
главные клетки
врожденного
иммунитета-
ФАГОЦИТЫ-
микрофагоциты
макрофагоциты
НГ
МОН/МФ
Фагоцитоз- особый процесс поглощения клеткой ,крупных макромолекулярных комплексов
главные клетки
врожденного
иммунитета-
ФАГОЦИТЫ-
микрофагоциты
макрофагоциты
НГ
МОН/МФ
Фагоцитоз- особый процесс поглощения клеткой ,крупных макромолекулярных комплексов

ФАГОЦИТОЗ
–комплекс клеточных событий, в основе которых лежит распознавание, поглощение и элиминация
ФАГОЦИТОЗ
–комплекс клеточных событий, в основе которых лежит распознавание, поглощение и элиминация

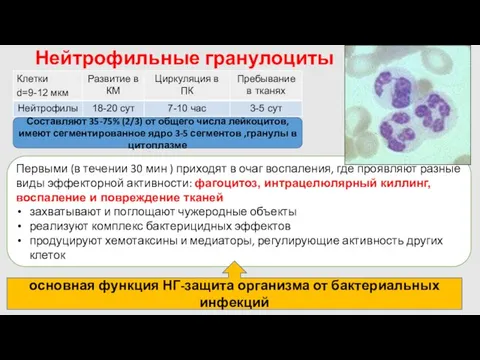
Нейтрофильные гранулоциты
Первыми (в течении 30 мин ) приходят в очаг воспаления,
Нейтрофильные гранулоциты
Первыми (в течении 30 мин ) приходят в очаг воспаления,

ПОВЕРХНОСТНЫЕ МЕМБРАННЫЕ РЕЦЕПТОРЫ НЕЙТРОФИЛЬНЫХ ГРАНУЛОЦИТОВ
CR3(CD11b)
CR1(CD35)
C5aR
LFA-1(CD11a)
VLA-4(CD49d)
FcγRIII (CD16)
CD18
FcγRIIA(CD32)
FcγRI(CD64)*
CR4(CD11c)
L-селектин(CD62L)
PSGL-1(CD162)
CD15
CD17
TLR1,2,4-10
CD14
FcαR
FcεR
ICAM-3(CD50)
CD40
MHC I
MHC II*
*- активированный НГ
CD95*
многочисленные рецепторы
ПОВЕРХНОСТНЫЕ МЕМБРАННЫЕ РЕЦЕПТОРЫ НЕЙТРОФИЛЬНЫХ ГРАНУЛОЦИТОВ
CR3(CD11b)
CR1(CD35)
C5aR
LFA-1(CD11a)
VLA-4(CD49d)
FcγRIII (CD16)
CD18
FcγRIIA(CD32)
FcγRI(CD64)*
CR4(CD11c)
L-селектин(CD62L)
PSGL-1(CD162)
CD15
CD17
TLR1,2,4-10
CD14
FcαR
FcεR
ICAM-3(CD50)
CD40
MHC I
MHC II*
*- активированный НГ
CD95*
многочисленные рецепторы
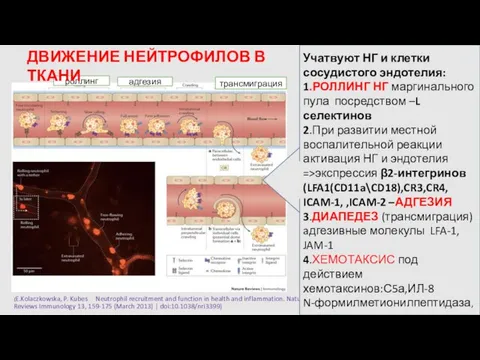
(E.Kolaczkowska, P. Kubes Neutrophil recruitment and function in health and inflammation.
(E.Kolaczkowska, P. Kubes Neutrophil recruitment and function in health and inflammation.

Распознавание чужеродных объектов НГ
основано на рецепторном взаимодействии фагоцита и бактериальной клетки
С
Распознавание чужеродных объектов НГ
основано на рецепторном взаимодействии фагоцита и бактериальной клетки
С

Поглощение и уничтожение микробов
Взаимодействие НГ с опсонизируемой частицей инициирует процесс поглощения
Поглощение и уничтожение микробов
Взаимодействие НГ с опсонизируемой частицей инициирует процесс поглощения
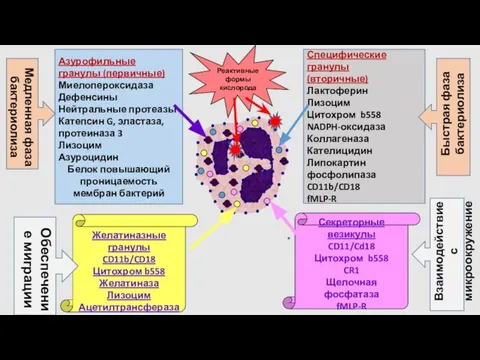
Специфические гранулы
(вторичные)
Лактоферин
Лизоцим
Цитохром b558
NADPH-оксидаза
Коллагеназа
Кателицидин
Липокартин
фосфолипаза
CD11b/CD18
fMLP-R
Азурофильные гранулы (первичные)
Миелопероксидаза
Дефенсины
Нейтральные протеазы
Катепсин G, эластаза, протеиназа 3
Лизоцим
Азуроцидин
Белок повышающий
Специфические гранулы
(вторичные)
Лактоферин
Лизоцим
Цитохром b558
NADPH-оксидаза
Коллагеназа
Кателицидин
Липокартин
фосфолипаза
CD11b/CD18
fMLP-R
Азурофильные гранулы (первичные)
Миелопероксидаза
Дефенсины
Нейтральные протеазы
Катепсин G, эластаза, протеиназа 3
Лизоцим
Азуроцидин
Белок повышающий
Механизмы разрушения чужеродных веществ
Кислородзависимый
Кислороднезависимый
гидролитические ферменты:
протеиназы,
катионные белки,
лизоцим,
Лактоферин
(активно связывает
Механизмы разрушения чужеродных веществ
Кислородзависимый
Кислороднезависимый
гидролитические ферменты:
протеиназы,
катионные белки,
лизоцим,
Лактоферин
(активно связывает

Кислород независимые факторы защиты
Естественные полипептидные антибиотики-
против Грамм(+), Грамм(-) бактерий, грибков и
Кислород независимые факторы защиты
Естественные полипептидные антибиотики-
против Грамм(+), Грамм(-) бактерий, грибков и
Кислород независимые факторы защиты
Серпроцидины-семейство сериновых нейтральных протеаз:
Эластаза
Азуроцидин- работает как ВPI-бактерицидный протеин
Протеиназа
Кислород независимые факторы защиты
Серпроцидины-семейство сериновых нейтральных протеаз:
Эластаза
Азуроцидин- работает как ВPI-бактерицидный протеин
Протеиназа

Природные антибиотики-связующее звено между врожденным и адаптивным иммунитетом
Дефензины НГ
индуцируют миграцию МОН,
Природные антибиотики-связующее звено между врожденным и адаптивным иммунитетом
Дефензины НГ
индуцируют миграцию МОН,

Кислород независимые факторы защиты
Лизоцим-(азурофильные и специфические гранулы НГ)
расщепляет пиптидогликаны микробной клетки,
Кислород независимые факторы защиты
Лизоцим-(азурофильные и специфические гранулы НГ)
расщепляет пиптидогликаны микробной клетки,

Инициация фагоцитоза
Запуск через
Поступающий в клетку О2 не используется для энергии
расходуется
Инициация фагоцитоза
Запуск через
Поступающий в клетку О2 не используется для энергии
расходуется

Респираторный взрыв
Кислород- зависимая реакция-миелопероксидаза независимая
Респираторный взрыв
Кислород- зависимая реакция-миелопероксидаза независимая

Респираторный взрыв
Кислород-зависимая реакция с участием Миелопероксидазы (МП)-фермент азурофильных гранул, основная функция
Респираторный взрыв
Кислород-зависимая реакция с участием Миелопероксидазы (МП)-фермент азурофильных гранул, основная функция
Цитохимические методы диагностики внутриклеточных ферментов клеток гранулоцитарного ряда
Положительная реакция
на
Цитохимические методы диагностики внутриклеточных ферментов клеток гранулоцитарного ряда
Положительная реакция
на
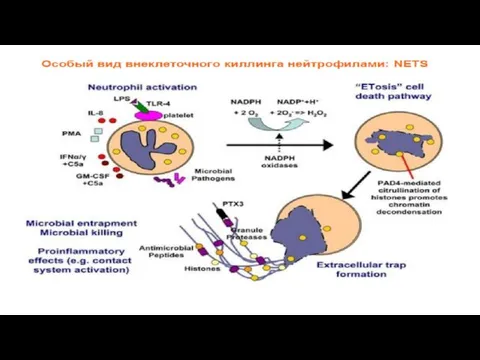
Экстрацеллюлярные нейтрофильные сети
(Neutrophil extracellular traps)
Активированные НГ экстрацеллюлярно выделяют белки гранул, ядерный
Экстрацеллюлярные нейтрофильные сети
(Neutrophil extracellular traps)
Активированные НГ экстрацеллюлярно выделяют белки гранул, ядерный
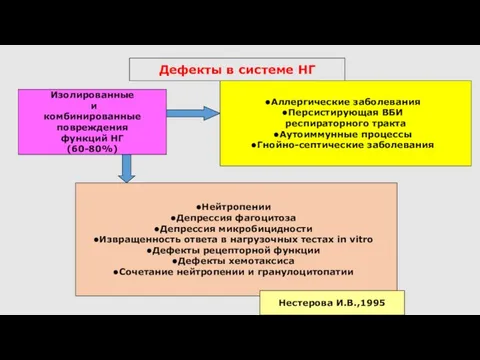
Дефекты в системе НГ
Аллергические заболевания
Персистирующая ВБИ
респираторного тракта
Аутоиммунные процессы
Гнойно-септические заболевания
Изолированные
и
Дефекты в системе НГ
Аллергические заболевания
Персистирующая ВБИ
респираторного тракта
Аутоиммунные процессы
Гнойно-септические заболевания
Изолированные
и

Макрофаги-ключевые клетки врожденного иммунитета
Способны распознавать, поглощать и убивать инфицирующие агенты
Очищать организм
Макрофаги-ключевые клетки врожденного иммунитета
Способны распознавать, поглощать и убивать инфицирующие агенты
Очищать организм

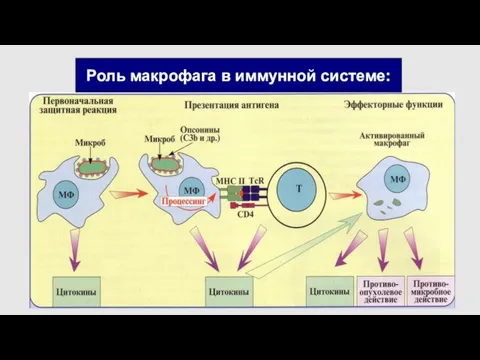
Роль макрофага в иммунной системе:
Роль макрофага в иммунной системе:

Основная функция МФ-очищение организма от умирающих клеток-имеет выраженный противовоспалительный характер
маркер CD14
scavenger
Основная функция МФ-очищение организма от умирающих клеток-имеет выраженный противовоспалительный характер
маркер CD14
scavenger

Продукты секреции макрофагов
Продукты секреции макрофагов

Различают МФ-резидентные и воспалительные
Резидентные возникают при «плановой» миграции в ткани без
Различают МФ-резидентные и воспалительные
Резидентные возникают при «плановой» миграции в ткани без

Миграционная и хемотаксическая функция МФ такая же как у НГ
хемоаттрактанты
фрагменты системы
Миграционная и хемотаксическая функция МФ такая же как у НГ
хемоаттрактанты
фрагменты системы
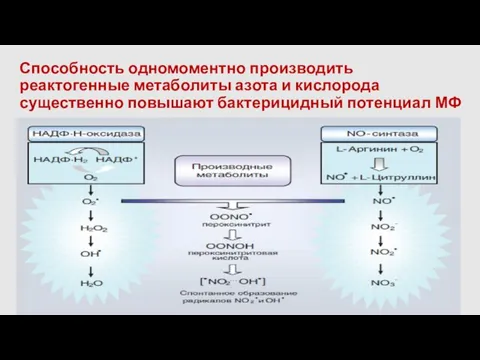
Способность одномоментно производить реактогенные метаболиты азота и кислорода существенно повышают бактерицидный
Способность одномоментно производить реактогенные метаболиты азота и кислорода существенно повышают бактерицидный
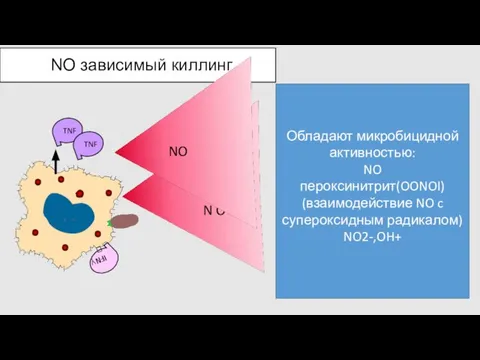
NO зависимый киллинг
Обладают микробицидной активностью:
NO
пероксинитрит(OONOI)
(взаимодействие NO c супероксидным радикалом)
NO2-,OH+
NO зависимый киллинг
Обладают микробицидной активностью:
NO
пероксинитрит(OONOI)
(взаимодействие NO c супероксидным радикалом)
NO2-,OH+

Дендритные клетки-профессиональные АПК
Превосходят МФ по экспрессии HLA II класса
Имеют костномозговое происхождение
В
Дендритные клетки-профессиональные АПК
Превосходят МФ по экспрессии HLA II класса
Имеют костномозговое происхождение
В

ЭОЗИНОФИЛЫ, ТУЧНЫЕ КЛЕТКИ ,БАЗОФИЛЫ
Основные эффекторы антипаразитарного иммунитета и аллергических реакций-открыл Пауль
ЭОЗИНОФИЛЫ, ТУЧНЫЕ КЛЕТКИ ,БАЗОФИЛЫ
Основные эффекторы антипаразитарного иммунитета и аллергических реакций-открыл Пауль
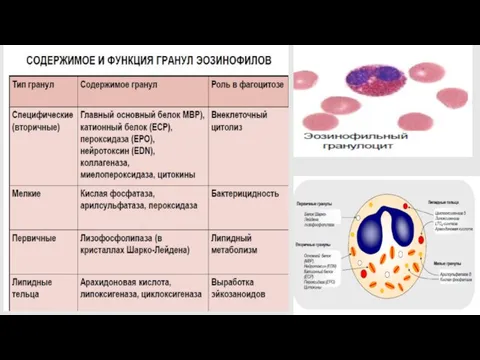
эозинофилы
эозинофилы
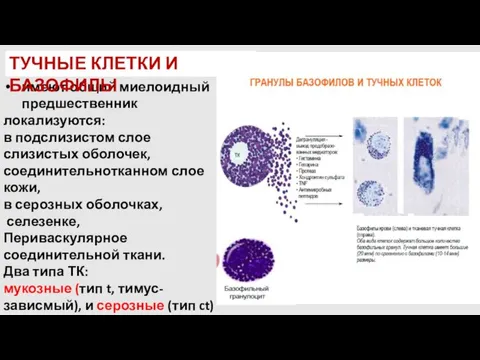
имеют общий миелоидный предшественник
локализуются:
в подслизистом слое слизистых оболочек, соединительнотканном слое
имеют общий миелоидный предшественник
локализуются:
в подслизистом слое слизистых оболочек, соединительнотканном слое
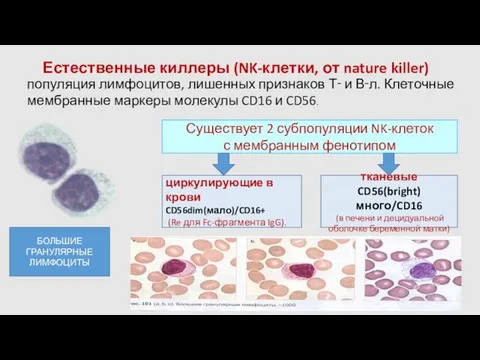
Естественные киллеры (NK-клетки, от nature killer)
циркулирующие в крови CD56dim(мало)/CD16+
(Re для
Естественные киллеры (NK-клетки, от nature killer)
циркулирующие в крови CD56dim(мало)/CD16+
(Re для
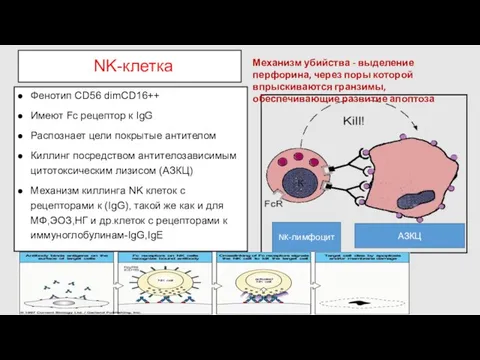
NK-клетка
Фенотип CD56 dimСD16++
Имеют Fc рецептор к IgG
Распознает цели покрытые антителом
Киллинг посредством
NK-клетка
Фенотип CD56 dimСD16++
Имеют Fc рецептор к IgG
Распознает цели покрытые антителом
Киллинг посредством

Активированные NK клетки цитокинами
IL2
IFN
IFN
IL2
Главная физиологическая функция – киллерная в отношении активированных
Активированные NK клетки цитокинами
IL2
IFN
IFN
IL2
Главная физиологическая функция – киллерная в отношении активированных

Другие рецепторы естественных киллеров
сложная система рецепторов распознающих
-молекулы собственных клеток организма
Другие рецепторы естественных киллеров
сложная система рецепторов распознающих
-молекулы собственных клеток организма

Активирующий и ингибирующий сигналы NK.
система регуляторных Re (inhibitory NK cell receptors).
2
Активирующий и ингибирующий сигналы NK.
система регуляторных Re (inhibitory NK cell receptors).
2

NKT - лимфоциты
«Неклассическая» субпопуляция Т-лимфоцитов врожденного иммунитета
сочетает признаки Т-л и NK,
NKT - лимфоциты
«Неклассическая» субпопуляция Т-лимфоцитов врожденного иммунитета
сочетает признаки Т-л и NK,

yδT – лимфоциты
В ранний период после рождения, выполняют важные иммунорегуляторные функции
yδT – лимфоциты
В ранний период после рождения, выполняют важные иммунорегуляторные функции

Часть 2.ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА
Часть 2.ГУМОРАЛЬНЫЕ ФАКТОРЫ ВРОЖДЕННОГО ИММУНИТЕТА

СИСТЕМА КОМПЛЕМЕНТА (С)-
-КОМПЛЕКС белков комплемента, их расщепленных фрагментов, рецепторов к компонентам
СИСТЕМА КОМПЛЕМЕНТА (С)-
-КОМПЛЕКС белков комплемента, их расщепленных фрагментов, рецепторов к компонентам

Белки системы комплемента
представлены сывороточными белками (> 30), циркулирующими в кровеносном русле
Белки системы комплемента
представлены сывороточными белками (> 30), циркулирующими в кровеносном русле

КОМПОНЕНТЫ КОМПЛЕМЕНТА СИНТЕЗИРУЮТСЯ:
в печени- гепатоцитами (90%)
макрофагами
эпителием кишечника, почечных канальцев
КОМПОНЕНТЫ КОМПЛЕМЕНТА СИНТЕЗИРУЮТСЯ:
в печени- гепатоцитами (90%)
макрофагами
эпителием кишечника, почечных канальцев
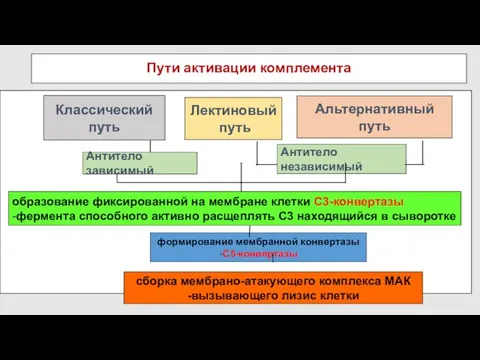
Пути активации комплемента
Классический путь
Альтернативный путь
Лектиновый
путь
Пути активации комплемента
Классический путь
Альтернативный путь
Лектиновый
путь
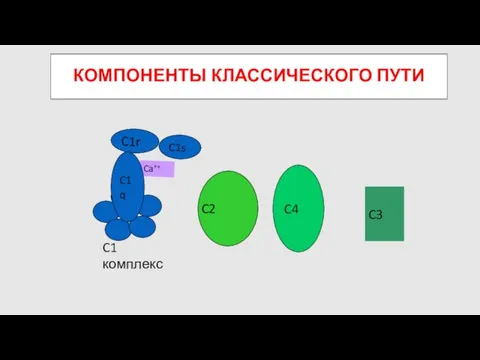
КОМПОНЕНТЫ КЛАССИЧЕСКОГО ПУТИ
C4
C2
C3
C1 комплекс
КОМПОНЕНТЫ КЛАССИЧЕСКОГО ПУТИ
C4
C2
C3
C1 комплекс

C4
КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ
C3-КОНВЕРТАЗЫ
Активация системы комплемента по классическому пути- индуцируется
C4
КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ
C3-КОНВЕРТАЗЫ
Активация системы комплемента по классическому пути- индуцируется
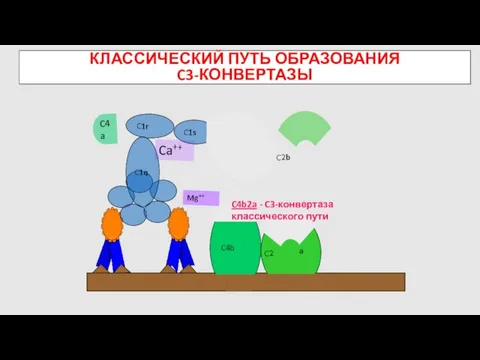
КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ
C3-КОНВЕРТАЗЫ
C4b
Mg++
C4a
C2
C4b2a - C3-конвертаза классического пути
КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ
C3-КОНВЕРТАЗЫ
C4b
Mg++
C4a
C2
C4b2a - C3-конвертаза классического пути
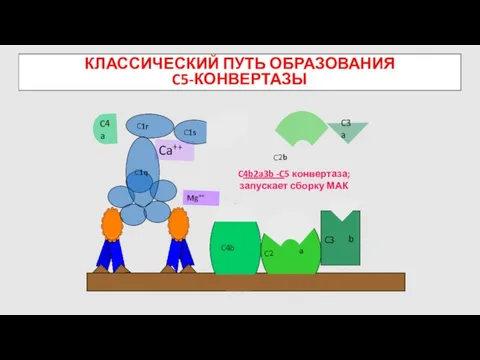
КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ
C5-КОНВЕРТАЗЫ
C4b
Mg++
C4a
C2b
C3
C4b2a3b -C5 конвертаза; запускает сборку МАК
КЛАССИЧЕСКИЙ ПУТЬ ОБРАЗОВАНИЯ
C5-КОНВЕРТАЗЫ
C4b
Mg++
C4a
C2b
C3
C4b2a3b -C5 конвертаза; запускает сборку МАК

Компоненты литического комплекса
C6
C9
C5
C9
C9
C9
Компоненты литического комплекса
C6
C9
C5
C9
C9
C9
Активация C5
C5
Активация C5
C5

Сборка литического комплекса МАК
C6
Сборка литического комплекса МАК
C6
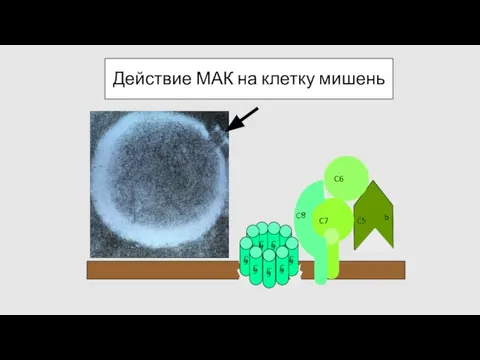
Действие МАК на клетку мишень
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
Действие МАК на клетку мишень
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9

Лектиновый путь
C4
C2
C4b2a - C3 конвертаза; активирует образование C5 -конвертазы
MASP1
MASP2
Активация -индуцируется манонозо-связывающим
Лектиновый путь
C4
C2
C4b2a - C3 конвертаза; активирует образование C5 -конвертазы
MASP1
MASP2
Активация -индуцируется манонозо-связывающим
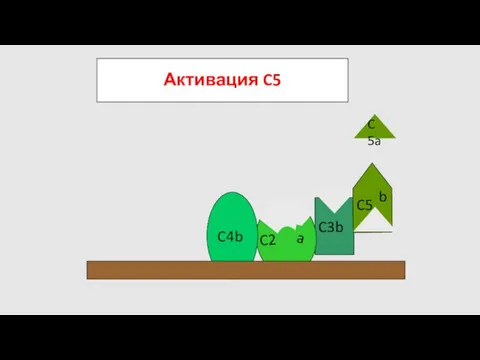
Активация C5
C5
Активация C5
C5

Сборка литического комплекса МАК
C6
Сборка литического комплекса МАК
C6
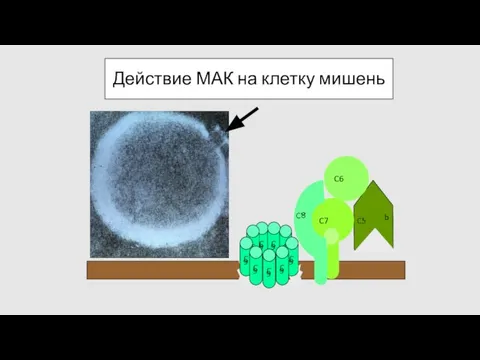
Действие МАК на клетку мишень
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
Действие МАК на клетку мишень
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9

Компоненты альтернативного пути
C3
B
D
P
Запуск начинается с С3
Содержание С3 составляет ½ концентрации всех
Компоненты альтернативного пути
C3
B
D
P
Запуск начинается с С3
Содержание С3 составляет ½ концентрации всех
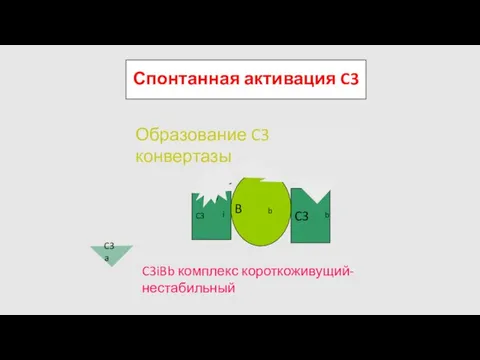
Спонтанная активация C3
C3
H2O
B
D
Образование C3 конвертазы
C3iBb комплекс короткоживущий-нестабильный
C3
Спонтанная активация C3
C3
H2O
B
D
Образование C3 конвертазы
C3iBb комплекс короткоживущий-нестабильный
C3

B
D
Спонтанное -образование C3b не разрушается
C3-активация и амплификация
C3
Комплекс C3bBb очень не
B
D
Спонтанное -образование C3b не разрушается
C3-активация и амплификация
C3
Комплекс C3bBb очень не
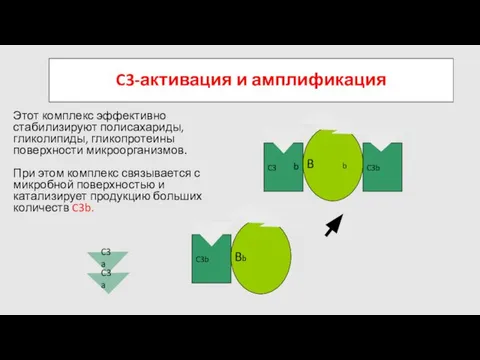
C3a
B
D
C3
C3-активация и амплификация
Этот комплекс эффективно стабилизируют полисахариды, гликолипиды, гликопротеины поверхности микроорганизмов.
C3a
B
D
C3
C3-активация и амплификация
Этот комплекс эффективно стабилизируют полисахариды, гликолипиды, гликопротеины поверхности микроорганизмов.
C3a
C3a
C3
B
D
C3-активация и амплификация
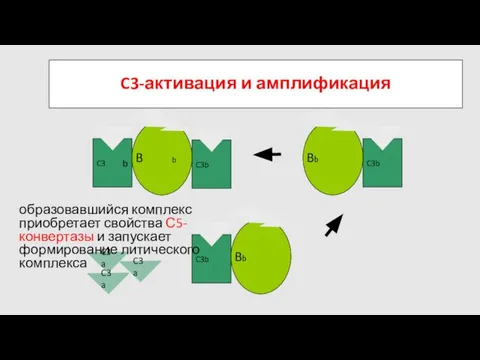
образовавшийся комплекс приобретает свойства С5-конвертазы и запускает формирование литического
C3a
C3a
C3
B
D
C3-активация и амплификация
образовавшийся комплекс приобретает свойства С5-конвертазы и запускает формирование литического
Активация C5
C5
Активация C5
C5

Сборка литического комплекса МАК
C6
Сборка литического комплекса МАК
C6
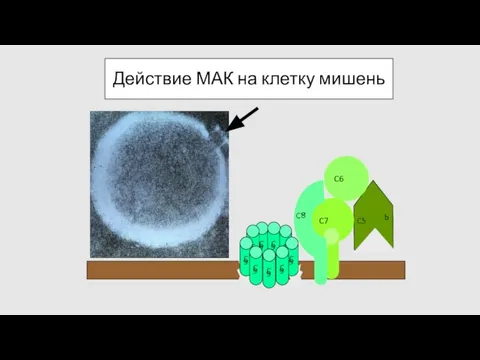
Действие МАК на клетку мишень
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
Действие МАК на клетку мишень
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9
C
9

C5-конвертазы классического, лектинового и альтернативного путей
C3b
Bb
C3b
C5-конвертаза
альтернативного пути
C5-конвертаза
классического и лектинового
C5-конвертазы классического, лектинового и альтернативного путей
C3b
Bb
C3b
C5-конвертаза
альтернативного пути
C5-конвертаза
классического и лектинового
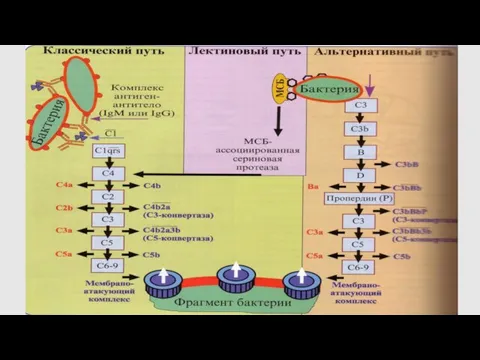

Центральный этап каскада комплемента
Все три пути активации С приводят к образованию
Центральный этап каскада комплемента
Все три пути активации С приводят к образованию


C3b
Опсонин- способствуют фагоцитозу бактерий,
на поверхности которых идет активация системы
C3b
Опсонин- способствуют фагоцитозу бактерий,
на поверхности которых идет активация системы
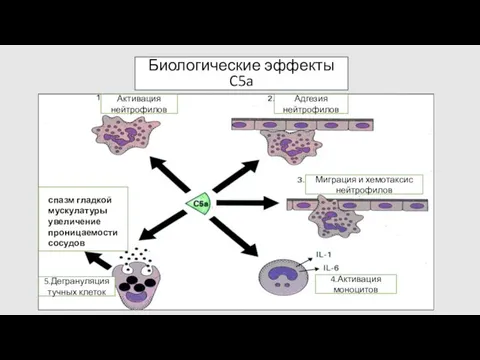
Биологические эффекты C5a
Активация нейтрофилов
Адгезия
нейтрофилов
Миграция и хемотаксис
нейтрофилов
4.Активация моноцитов
5.Дегрануляция тучных клеток
спазм гладкой
Биологические эффекты C5a
Активация нейтрофилов
Адгезия
нейтрофилов
Миграция и хемотаксис
нейтрофилов
4.Активация моноцитов
5.Дегрануляция тучных клеток
спазм гладкой
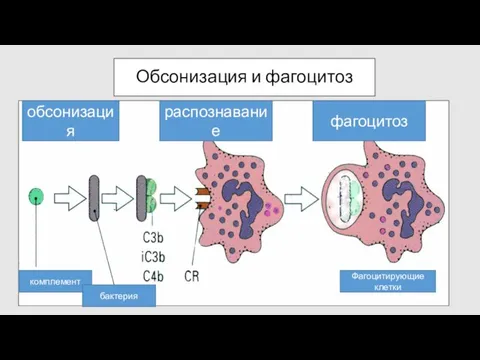
Обсонизация и фагоцитоз
комплемент
бактерия
Фагоцитирующие клетки
обсонизация
распознавание
фагоцитоз
Обсонизация и фагоцитоз
комплемент
бактерия
Фагоцитирующие клетки
обсонизация
распознавание
фагоцитоз
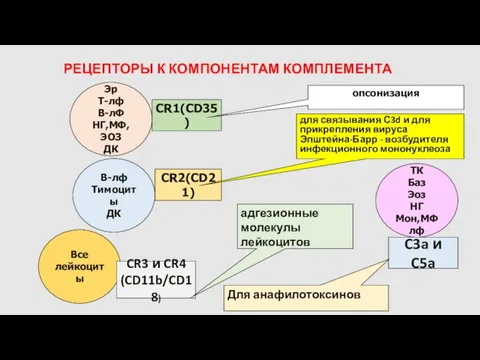
РЕЦЕПТОРЫ К КОМПОНЕНТАМ КОМПЛЕМЕНТА
Эр
Т-лф
В-лФ
НГ,МФ,ЭОЗ
ДК
CR1(СD35)
опсонизация
В-лф
Тимоциты
ДК
CR2(СD21)
для связывания С3d и для прикрепления вируса Эпштейна-Барр
РЕЦЕПТОРЫ К КОМПОНЕНТАМ КОМПЛЕМЕНТА
Эр
Т-лф
В-лФ
НГ,МФ,ЭОЗ
ДК
CR1(СD35)
опсонизация
В-лф
Тимоциты
ДК
CR2(СD21)
для связывания С3d и для прикрепления вируса Эпштейна-Барр

РЕГУЛЯЦИЯ АКТИВНОСТИ КОМПЛЕМЕНТА
Регуляторы – белки, подавляющие отдельные стадии каскада активации С,
РЕГУЛЯЦИЯ АКТИВНОСТИ КОМПЛЕМЕНТА
Регуляторы – белки, подавляющие отдельные стадии каскада активации С,

РЕГУЛЯЦИЯ АКТИВНОСТИ КОМПЛЕМЕНТА
Растворимые:
Ингибитор С1 - эстеразы (C1-INH или С1-inh) блокирует протеолиз
РЕГУЛЯЦИЯ АКТИВНОСТИ КОМПЛЕМЕНТА
Растворимые:
Ингибитор С1 - эстеразы (C1-INH или С1-inh) блокирует протеолиз

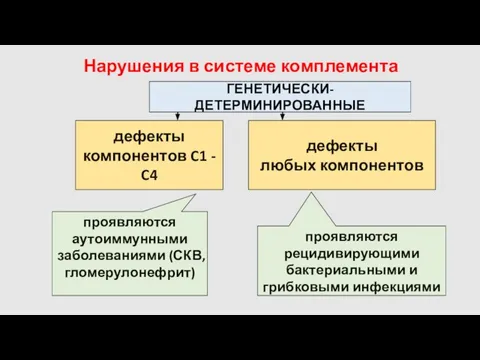
Нарушения в системе комплемента
ГЕНЕТИЧЕСКИ-ДЕТЕРМИНИРОВАННЫЕ
дефекты
компонентов C1 - C4
дефекты
любых компонентов
проявляются аутоиммунными
Нарушения в системе комплемента
ГЕНЕТИЧЕСКИ-ДЕТЕРМИНИРОВАННЫЕ
дефекты
компонентов C1 - C4
дефекты
любых компонентов
проявляются аутоиммунными
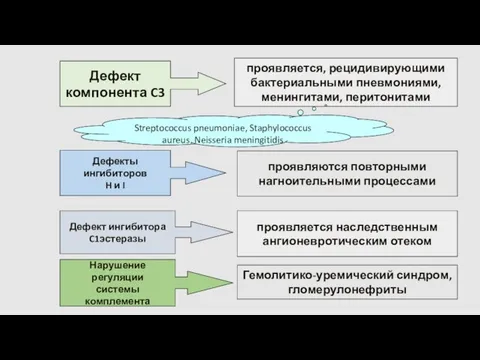
Дефект
компонента C3
проявляется, рецидивирующими
бактериальными пневмониями,
менингитами, перитонитами
Streptococcus pneumoniae, Staphylococcus aureus, Neisseria
Дефект
компонента C3
проявляется, рецидивирующими
бактериальными пневмониями,
менингитами, перитонитами
Streptococcus pneumoniae, Staphylococcus aureus, Neisseria
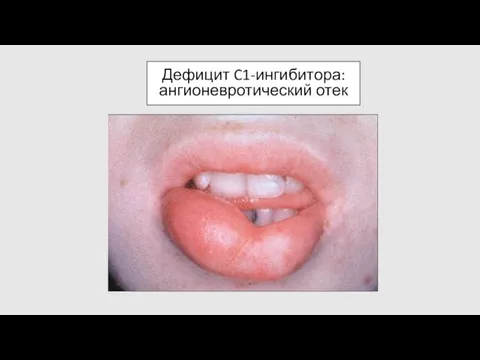
Дефицит C1-ингибитора: ангионевротический отек
Дефицит C1-ингибитора: ангионевротический отек
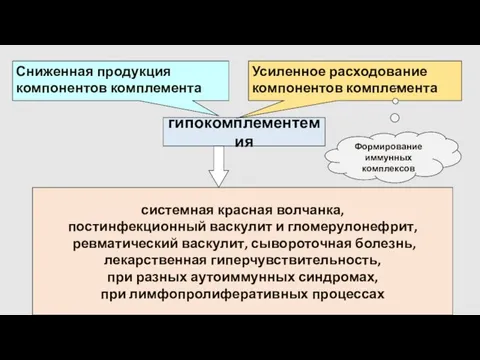
гипокомплементемия
Сниженная продукция компонентов комплемента
Усиленное расходование
компонентов комплемента
Формирование
иммунных комплексов
системная красная волчанка,
постинфекционный васкулит
гипокомплементемия
Сниженная продукция компонентов комплемента
Усиленное расходование
компонентов комплемента
Формирование
иммунных комплексов
системная красная волчанка,
постинфекционный васкулит

-группа протеинов синтезируемая гепатоцитами.
В ответ на стрессорные факторы (воспаление, малигнизация,
В ответ на стрессорные факторы (воспаление, малигнизация,
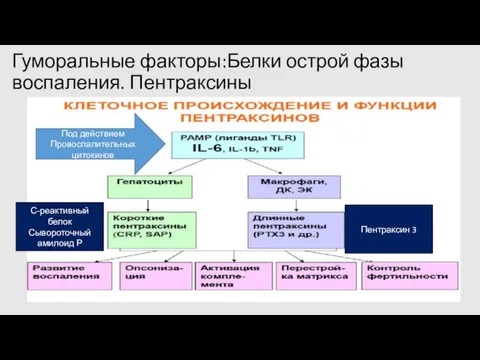
Гуморальные факторы:Белки острой фазы воспаления. Пентраксины
С-реактивный белок
Сывороточный амилоид Р
Под действием
Провоспалительных цитокинов
Пентраксин
Гуморальные факторы:Белки острой фазы воспаления. Пентраксины
С-реактивный белок
Сывороточный амилоид Р
Под действием
Провоспалительных цитокинов
Пентраксин

С- реактивный белок (СРБ)
В норме (≈ 1 мкг/мл), при воспалительных
С- реактивный белок (СРБ)
В норме (≈ 1 мкг/мл), при воспалительных

Маннозосвязывающий лектин – МСЛ (MBL)- относится к семейству коллектинов -
Маннозосвязывающий лектин – МСЛ (MBL)- относится к семейству коллектинов -
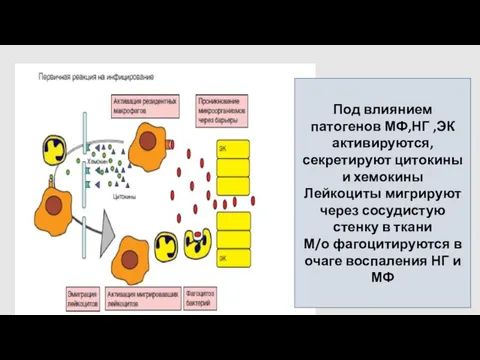
Под влиянием патогенов МФ,НГ ,ЭК активируются, секретируют цитокины и хемокины
Лейкоциты мигрируют
Под влиянием патогенов МФ,НГ ,ЭК активируются, секретируют цитокины и хемокины
Лейкоциты мигрируют

Фагоциты. В начале данного этапа их эффективность невелика:
число НГ и МФ
число НГ и МФ
 Вирусы
Вирусы Потребность пациента в поддержании нормальной температуры тела
Потребность пациента в поддержании нормальной температуры тела Острая артериальная ишемия
Острая артериальная ишемия Синтетические противомикробные средства
Синтетические противомикробные средства Внезапные состояния при заболеваниях желудочно-кишечного тракта, эндокринной системы и органов мочевыделения. (Лекция 6)
Внезапные состояния при заболеваниях желудочно-кишечного тракта, эндокринной системы и органов мочевыделения. (Лекция 6) Тромболизис. Тромболитическая терапия
Тромболизис. Тромболитическая терапия Еркек жыныс мүшесін қалыңдату әдістері
Еркек жыныс мүшесін қалыңдату әдістері Черепно-мозговая травма. Первая медицинская и доврачебная помощь
Черепно-мозговая травма. Первая медицинская и доврачебная помощь Опухолевые стволовые клетки на примере рака молочной железы
Опухолевые стволовые клетки на примере рака молочной железы Болевые синдромы поясничной области и их коррекция
Болевые синдромы поясничной области и их коррекция Анемия. Виды анемий
Анемия. Виды анемий Тулеремия және оба қоздырғыштары
Тулеремия және оба қоздырғыштары Дыхательная недостаточность
Дыхательная недостаточность Phylogenetic disorders of skull
Phylogenetic disorders of skull Кровотечения в последовом и в раннем послеродовом периоде
Кровотечения в последовом и в раннем послеродовом периоде Заболевания придатков глаза, роговицы и склеры. Лекция №2
Заболевания придатков глаза, роговицы и склеры. Лекция №2 Өлімнің коммуникативтік этикалық мәселелері
Өлімнің коммуникативтік этикалық мәселелері Кожа – индикатор здоровья
Кожа – индикатор здоровья Актуальные вопросы радиационной гигиены в отечественной медицине
Актуальные вопросы радиационной гигиены в отечественной медицине Показатели детской инвалидности муз дгб№4
Показатели детской инвалидности муз дгб№4 Рекомендации по созданию презентации на научно-практические конференции по медицине
Рекомендации по созданию презентации на научно-практические конференции по медицине Рганизация всех видов медицинских осмотров. Медсанчасти, медпункты, здравпункты предприятий. Производственная аптечка
Рганизация всех видов медицинских осмотров. Медсанчасти, медпункты, здравпункты предприятий. Производственная аптечка Сестринская деятельность в реабилитации пациентов с патологией ЦНС
Сестринская деятельность в реабилитации пациентов с патологией ЦНС Подагра
Подагра Клиническое обследование. Clinical examination. Лекция 2
Клиническое обследование. Clinical examination. Лекция 2 Меры предотвращения распространения гриппа, острых респираторных вирусных инфекций и новой коронавирусной инфекции
Меры предотвращения распространения гриппа, острых респираторных вирусных инфекций и новой коронавирусной инфекции Бактерийные и вирусные лечебные, профилактические и диагностические препараты
Бактерийные и вирусные лечебные, профилактические и диагностические препараты Офтальмоскопические признаки диабетической ретинопатии
Офтальмоскопические признаки диабетической ретинопатии